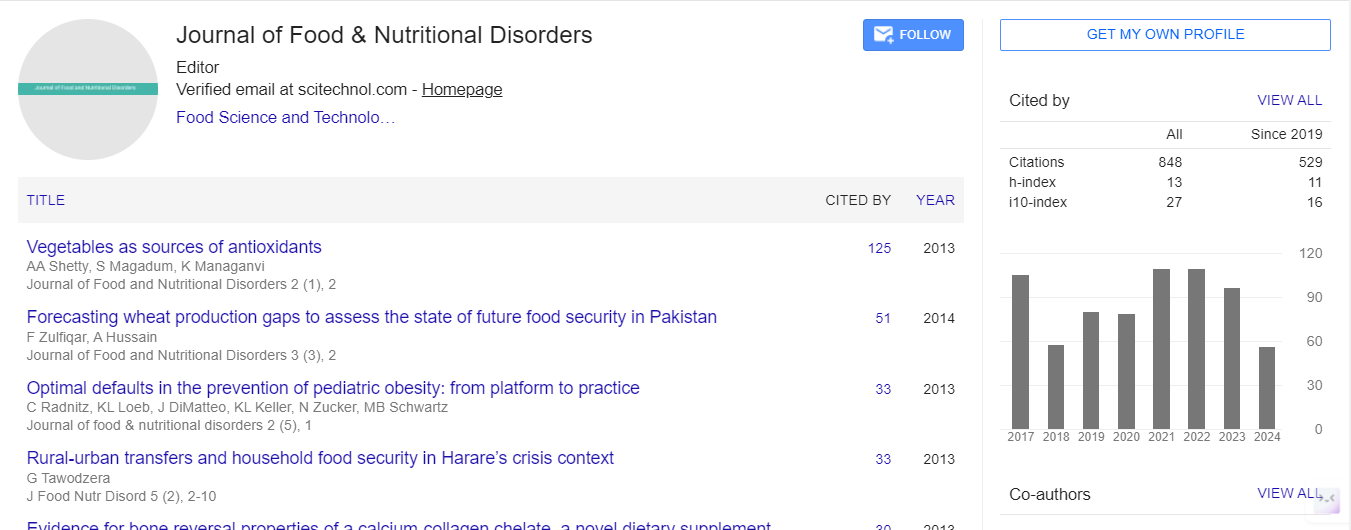
Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Editorial, J Food Nutr Disor Vol: 4 Issue: 1
What's bugging you and your diet?
| Bryn L Yeomans* and Nicholas T Bello | |
| Department of Animal Sciences, New Jersey Institute for Food, Nutrition and Health, Rutgers, The State University of New Jersey, New Brunswick, NJ 08901, USA | |
| Corresponding author : Bryn L Yeomans Department of Animal Sciences, Nutritional Science Graduate Program, New Jersey, USA E-mail: bly8@scarletmail.Rutgers.edu |
|
| Received: January 25, 2015 Accepted: January 27, 2015 Published: January 30, 2015 | |
| Citation: Yeomans BL, Bello NT (2015) What’s Bugging you and your Diet? J Food Nutr Disor 4:1. doi:10.4172/2324-9323.1000e110 |
Abstract
What’s Bugging you and your Diet?
Trillions of microorganisms live in and on our bodies. These are active communities of bacteria, viruses, archaea, and unicellular eukaryotes, collectively known as the microbiota. These organisms are involved in the development and regulation of host immunity, digestion and absorption, metabolism, behavior, and protection from pathogens. The microbiota inhabits all external surfaces of the body. These include the skin, and the urogenital, respiratory, and gastrointestinal tracts, with an estimated 70% of the microbiota residing in the colon . While the human gut microbiota is comprised of more than 50 bacterial phyla, the two that predominate are Bacteroidetes and Firmicutes, with Proteobacteria, Verrucomicrobia, Actinobacteria, Fusobacteria, and Cyanobacteria having a lesser presence.
| Trillions of microorganisms live in and on our bodies. These are active communities of bacteria, viruses, archaea, and unicellular eukaryotes, collectively known as the microbiota. These organisms are involved in the development and regulation of host immunity, digestion and absorption, metabolism, behavior, and protection from pathogens. The microbiota inhabits all external surfaces of the body. These include the skin, and the urogenital, respiratory, and gastrointestinal tracts, with an estimated 70% of the microbiota residing in the colon [1]. While the human gut microbiota is comprised of more than 50 bacterial phyla, the two that predominate are Bacteroidetes and Firmicutes, with Proteobacteria, Verrucomicrobia, Actinobacteria, Fusobacteria, and Cyanobacteria having a lesser presence [2]. Humans display a high degree of interindividual variation at the genus and species level, with more than 35,000 bacterial species having been identified within the GI [3]. Furthermore, the density and diversity of the microbial composition along the length of the GI increases with passage from the proximal to distal regions [4], and also shows heterogeneity when comparing the lumen, mucus layer, and epithelial crypts [5]. | |
| Colonization of the human gut begins at birth, and differs with maternal microbial make-up as well as mode of delivery. Microbial communities remain relatively undeveloped until the age of one, and by 18-24 months of age [1], the gut microbiota is considered to be “adult-like” in stability and resilience when 3 challenged [6]. Microbiota composition is determined, therefore, by external factors such as maternal microbiota, and interaction between host genetics and age, diet, exposure to pathogens, and antibiotic use. Although stable in terms of the bacterial communities present, the gut microbiota is susceptible to changes in the relative amounts of representative microorganisms due to these external factors [7]. | |
| Host diet has an immediate and measurable effect on the gut microbiota. When switching humanized C57BL/6J mice from a lowfat/ high-plant polysaccharide diet to a high-fat/high-sugar “Western” diet, Turnbaugh and colleagues found alterations in microbial composition and microbiome gene expression after only one day [8]. Similar responses were found by Duca and colleagues that observed diet-dependent differences in the gut microbiota of obese-prone (OP) and obese-resistant (OR) rats on standard chow or high fat (HF) diet. While OP and OR rats showed no phylum level differences when fed a standard chow, the microbial profiles diverged after 12 weeks on a HF diet, and there were significant differences between HF-fed rats and their chow-fed counterparts [9]. Although the diet-microbiome connection has been illustrated by numerous studies [10-12], the relative contribution of diet compared to other environmental factors [13] and whether the findings extend to human dietary patterns is controversial [4]. | |
| The gut microbiota can be viewed as a polygenic trait shaped by numerous environmental and genetic factors [14,15]. Benson et al. performed quantitative 16S rDNApyrosequencing of the gut microbiota in a mouse advanced intercross line (AIL) to systematically investigate the contribution of host genetics to microbial composition. A core measurable microbiota (CMM) of 64 taxonomic groups conserved across the animal population, but varied in quantity was identified, with litter, cohort, and individual host genetics contributing to composition. Quantitative trait loci (QTL) analysis was used to explore the contribution of host genotype to the variation in CMM in the AIL population; identifying 26 taxonomic groups associated with 13 significant QTL. Many QTL were found to have pleiotropic effects, with some acting on two phylogenetically similar organisms, some acting on organisms of different taxonomic levels, and others acting similarly on phylogenetically distant microbes. | |
| In order explore the genetic contribution to the gut microbiota in an obesogenic environment, Parks and colleagues utilized 16S rRNApyrosequencing to analyze the cecal microbial content of more than 100 strains of inbred mice fed a high-fat/high-sucrose diet for 8 weeks (Parks et al., 2013)[8]. Their findings suggested that HF/HS feeding alters gut microbiota composition across a variety of genetic backgrounds. | |
| Along these lines, Lees and colleagues investigated a model of genetically-induced obesity to explore the relative contribution of host genetics, cage environment, and age on gut bacterial composition [16,17]. Homozygous Zucker (fa/fa) rats, which have a recessive mutation of the leptin receptor gene, develop hyperphagia, [5] Obesity, and insulin resistance. Homozygous obese (fa/fa), heterozygous lean (fa/+), and homozygous lean (+/+) Zucker rats were co-housed, and metabolic parameters were measured and fecal samples analyzed using 454 pyrosequencing at 5, 7, 10, and 14 weeks of age. Overall age-effects were found to dominate gut microbial differences. Cage-associated trends were seen when time points were considered independently. Surprisingly, no genotypic differences were detected suggesting differences between phenotype are a result of obesogenic feeding rather than host genetic background. | |
| It is clear that the development and maintenance of the gut microbiota reflects the interaction between maternal microbiota, genetics, housing, diet, pathogenic exposure, antibiotic use, and age. However, it is the intersection of these genetic, environmental, and stochastic factors that warrants continued research. What is the extent of the genetic contribution, and are there specific instances when the effect can be minimized by the presence of other factors? Due to the close interaction between our microbiota and our innate and adaptive immune systems, do functional polymorphisms in immune-related genes have an exaggerated effect? Are there genotypes that are differentially predisposed to obesity or disease when on a high-fat, high-sugar diet; and can we manipulate or attenuate this susceptibility via dietary intervention? Future research seeks to answer these questions, and the underlying query; how do we optimize our relationship with these commensal critters to promote improvements in human health and reduce disease susceptibility [6]? |
|
References
- Ley RE, Peterson DA, Gordon JI (2006) Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 124: 837-848.
- Frank DN, Amand AL, Feldman RA, Boedeker EC, Harpaz N, et al. (2007) Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. ProcNatlAcadSci USA 104: 13780-13785.
- Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, et al. (2005) Diversity of the human intestinal microbial flora. Science 308: 1635-1638.
- O'Hara AM, Shanahan F (2006) The gut flora as a forgotten organ. EMBO Rep 7: 688-693.
- Swidsinski A, Loening-Baucke V, Lochs H, Hale LP (2005) Spatial organization of bacterial flora in normal and inflamed intestine: a fluorescence in situ hybridization study in mice. World J Gastroenterol 11: 1131-1140.
- Antonopoulos DA, Huse SM, Morrison HG, Schmidt TM, Sogin ML, et al. (2009)Reproducible community dynamics of the gastrointestinal microbiota following antibiotic perturbation. Infect Immun, 6: 2367-2375.
- David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, et al. (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505: 559-563.
- Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, et al. (2009) The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. SciTransl Med 1: 6ra14.
- Duca FA, Sakar Y, Lepage P, Devime F, Langelier B, et al. (2014) Replication of obesity and associated signaling pathways through transfer of microbiota from obese-prone rats. Diabetes 63: 1624-1636.
- Zhang C, Zhang M, Pang X, Zhao Y, Wang L, et al. (2012) Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J 6: 1848-1857.
- Parks BW, Nam E, Org E, Kostem E, Norheim F, et al. (2013) Genetic control of obesity and gut microbiota composition in response to high-fat, high-sucrose diet in mice. Cell Metab 17: 141-152.
- Kashyap PC, Marcobal A, Ursell LK, Smits SA, Sonnenburg ED, et al. (2013) Genetically dictated change in host mucus carbohydrate landscape exerts a diet-dependent effect on the gut microbiota. ProcNatlAcadSci U S A 110: 17059-17064.
- Wu GD, Compher C, Chen EZ, Smith SA, Shah RD, et al. (2014) Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production Gut.
- Benson AK, Kelly SA, Legge R, Ma F, Low SJ, et al. (2010) Individuality in gut microbiota composition is a complex polygenic trait shaped by multiple environmental and host genetic factors. ProcNatlAcadSci U S A 107: 18933-18938.
- McKnite AM, Perez-Munoz ME, Lu L, Williams EG, Brewer S, et al. (2012) Murine gut microbiota is defined by host genetics and modulates variation of metabolic traits. PLoS One 7: e39191.
- Lees H, Swann J, Poucher SM, Nicholson JK, Holmes E, et al. (2014) Age and microenvironment outweigh genetic influence on the Zucker rat microbiome. PLoS One 9: e100916.
- Carmody RN, Gerber GK, Luevano JM Jr, Gatti DM, Somes L, et al. (2015) Diet dominates host genotype in shaping the murine gut microbiota. Cell Host Microbe 17: 72-84.