Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, J Vet Sci Med Diagn Vol: 10 Issue: 7
Vitamin E in Vitro Action on Horse Red Blood Cell Membranes from Rest and Exercised Horses: Modulation of Membrane Fluidity and Glutathione Peroxidase Activity
Giuseppe Gallo* and Guglielmo Martino
Department of Biology, Ecology and Earth Sciences, University of Calabria, Rende CS, Italy
*Corresponding Author:Giuseppe Gallo
Department of Biology
Ecology and Earth Sciences
University of Calabria
Rende CS
Italy
E-mail: pino72@tiscali.it
Received: October 11, 2021; Accepted date: October 26, 2021; Published date: November 2, 2021
Citation: Gallo G, Martino G (2021) Vitamin E In Vitro Action on Horse Red Blood Cell Membranes from Rest and Exercised Horses: Modulation of Membrane Fluidity and Glutathione Peroxidase Activity. J Vet Sci Med Diagn 10:11.
Abstract
Objectives: The intensive exercise increases circulating radical oxygen specie levels in humans. Horses are athletic animals that can be trained to execute performances well established by International Equestrian Federations. They are useful models to study their physical performances and related structural and metabolic derangements.
Methods: The animals were ten Arabian strain horses, 6-8 years old, trained for Cross country and six horses at rest. Blood samples are tested for Glutathione Pero-Xidase (GPX) and chemical parameters both before and after standard exercise. Membranes are tested for diphenylhexatriene fluorescence anisotropy in red blood cell membranes by Perrin plot.
Results: Glutathione peroxidase activity determined in exercised animals and radical oxygen species at rest and preand post-exercise significantly decrease after exercise with P: 0.008 by Student-T. The vitamin addition (50 µM) to red blood cell membranes brings the activity to reference level. Fluorescence anisotropy is abruptly worsened in erythrocyte membranes from exercised animals and almost restored after vitamin E treatment of membrane preparations and P > 0.05 by ANOVA.
Discussion: Erythrocyte membranes are damaged by standard exercise and both GPX and membrane fluorescence anisotropy (rs) is almost restored to reference and pre-exercise levels by in vitro treatment with vitamin E.
Keywords: Horse; Vitamin E; Membrane fluidity; Red blood; cell membranes; Glutathione peroxidase; Oxidative stress; Physical exercise; Red blood cell indexes
Keywords
Horse; Vitamin E; Membrane fluidity; Red blood cell membranes; Glutathione peroxidase; Oxidative stress; Physical exercise; Red blood cell indexes
Introduction
Physical exercise increases oxidative stress and causes disruption of the energetic homeostasis [1]. In literature, experimental data also exist for physical exercise and different activities such as Glutathione Peroxidase (GPX), Red Blood Cell (RBC) activities of GPX and Total Antioxidant Status (TAS) [2-5]. Physical exercise in horses induces a series of normal physiological and biochemical adjustments [3]. Increasing metabolism and oxygen uptake may induce oxidative stress in various organs [4,5]. Associations between increased muscle leakage and decreased antioxidant status may, in part, reflect oxidative stress and suggest the testing of antioxidant supplements in endurance horses to improve performance and welfare. One study examined exercise-induced changes in some plasma and RBC biochemical and antioxidant variables only in pentathlon horses. Horses which were supplemented with vitamin E, at nearly 10-times the 1989 NRC recommended level, did not experience lower oxidative stress as compared to the control ones [6]. The relevance of antioxidant supplementation becomes apparent if the diet is deficient in antioxidants, if exercise intensity is higher or more prolonged, even if diseases or additional stresses are present. According to Avellini, the supplementation of vitamin E in a horse’s diet [7] enhances antioxidant defences in athletic horses, thus slowing down the peroxidation phenomena in exercised horse’s muscle and blood.
The aim of the present research is to detail GPX enzyme kinetic data and their alterations in trained animals, both before and after physical exercise in official competitions, and to evaluate ’In vitro’ the functional alterations of RBC membrane lipids (membrane fluidity), also under the action of standardized concentrations of vitamin E.
Materials and Methods
Animals
Horses are useful models to study their physical performances and related structural and metabolic derangements.
Two categories of horses were selected: horses at rest (6 animals) cross-country horses (10 animals).
16 male horses, 6-8 years old, Arabian strain, were used. 6 horses at rest and 10 trained for cross-country competitions and subjected to testing. In brief, the characteristics of the categories are the following: Horses at rest are subjects which are not trained and which are free of movement and without imposing physical exercise. Cross-country horses are trained 70 days, 6 days per week, with gradual increase up to race performance. The diet is as specified in Table 1.
| Ingredients | % (W/W) |
|---|---|
| Oats | 19.2 |
| Barley | 18.3 |
| Carob beans | 5 |
| Cottonseed meal | 6 |
| Limestone | 0.5 |
| Fescue hay | 51 |
| Total | 100 |
| Chemical composition | |
| Digestible energy Mcal/kg | 2.4 |
| Crude protein | 10.9 |
| Selenium (µg/kg) | 20 |
| Vitamin E (mg/kg) | 20-40 |
Table 1: Composition of horse diet, Vitamin E and Selenium added as supplements, Dry matter basis.
The race is in the open field for at least 2.5 km of race track (regional-national, Italian regularity F category).
The rations were supplemented with 20 µg/kg per day of selenium and 20-40 mg/kg per day of vitamin E.
Samples
The studied membrane model is the Red Blood Cell (RBC) that suffers damages from ROS [3]. The RBC is a useful experimental model of essential cellular structure. In particular, the equivalent reducing power deriving from the activity of the Glucose-6-Phosphate De-Hydrogenase (G6PDH) is the main source to feed the above mentioned scavenger. Exercised animal blood samples (5 ml) were collected 10 minutes before exercise, in vacutainers containing 0.15% EDTA and refrigerated at 8°C and after 5 minutes of recovery, post-exercise periods. All samples were drawn by aseptic jugular venipuncture performed by official vets and with the consent of horse owners. Samples were drawn from all horses by the upper one third of the jugular vein while occluding the vessel distally. 8 ml were transferred immediately into an evacuated, heparinized blood collection tube for chemical clinical analyses and 3 ml into an evacuated tube containing ethylene diaminetetraacetic acid were for GPX analyses.
All housing and care by owners were conforming to the standards recommended by the guide for the care and use of laboratory animals and directive 86/609 CEE. Experimentation was authorized by the Italian Ministry of Health (IMH), veterinary division, according to Italian ‘animal care’ (DL 116/92).
Glutathione peroxidase assay
Glutathione peroxidase, antioxidant enzyme (PDB 1GP1, EC 1.11.1.9), is selenium dependent.
GPX activity is determined according to Paglia et al [8] as modified by Gallo et al. [9-13] In RBC samples from the same animals, both before and after exercise (cross-country). The reagents were all analytical grade from Sigma (St. Louis USA).
In vitro α-tocopherol treatment
This scavenging activity was studied after the “In vitro” treatment with vitamin E, the most effective one. Vitamin E is the most commonly supplemented antioxidant in horses. Each sample contained 1 mg/mL α-tocopherol dissolved in Dimetyl Sulfoxide (DMSO) was added to reach a final concentration of 50, 100 or 150 µg α-tocopherol/mL plasma, that is similar to data of Williams et al. 2006 [5]. Tubes were incubated in a 37°C water bath under nitrogen, and in the dark, for 6 hrs. Samples were mixed by vortexing briefly every 30 minutes.
The α-tocopherol contents in serum of horse at rest and before and after exercise are respectively 2.33 ± 1.29 µg/ml, 2.16 ± 0.88 µg/ml and 1.77 ± 0.85 µg/ml.
Shinitzky spectrofluorometric method
The fluidity of membrane was determined by the shinitzky method [14-22]. which evaluates the parameter on a MPF 44B Perkin Elmer spectrofluorometer. The Fluorescence Polarization (FP) of a labeled macromolecule depends on the fluorescence lifetime (τ) and the rotational correlation time (θ)

Where P0 is the polarization observed in the absence of rotational diffusion as described in literature [19]. Polarization (P) ranges from –0.33 to +0.5 while the range for anisotropy (r) is from –0.25 to +0.4, where
P=3r/2+r
r=2P/3-p
DPH (diphenylhexatriene) makes its fluorescence polarization sensitive to binding interactions over a larger molecular weight range [23]. DPH in Tetrahydrofuran (THF), spectroscopy grade, into the buffer solution was used. Fluorescence polarization is measured by exciting the fluorophore with monochromatic light (~360 nm for DPH) through a polarizer whose axis is oriented vertically and parallel to the light path; the emission intensity is detected through an analyzer, whose polarization axis is oriented sequentially perpendicular. The emission is detected at 390 nm. The DPH solution in THF added to each sample is incubated 30 min at 40 °C before measurement.
The main haematological parameters are referred in Table 2.
| Indexes | Rest | C-C before | C-C after |
|---|---|---|---|
| Hb (g\100 ml) | 13.0 ± 0.60 | 12.4 ± 0.80 | 15.6 ± 0.90 |
| Ht (%) | 32.00 ± 0.53 | 33.50 ± 2.00 | 42.30 ± 2.60 |
| RBC count (E+06/µl) | 7.11E+06 ± 0.25 | 8.79E+06 ± 0.31 | 14.46E+06 ± 0.60 |
| MCV (µl) | 0.0000045 | 0.0000038 | 0.0000029 |
| Red blood cell polymorphisms | Anisocytosis | Poikilocytosis | Hypochromia |
Table 2: Main RBC indexes, each value are the mean (± SEM) of 6 independent determinations. Evaluation scale of RBC morphological heterogeneity of 5.
Statistical analysis
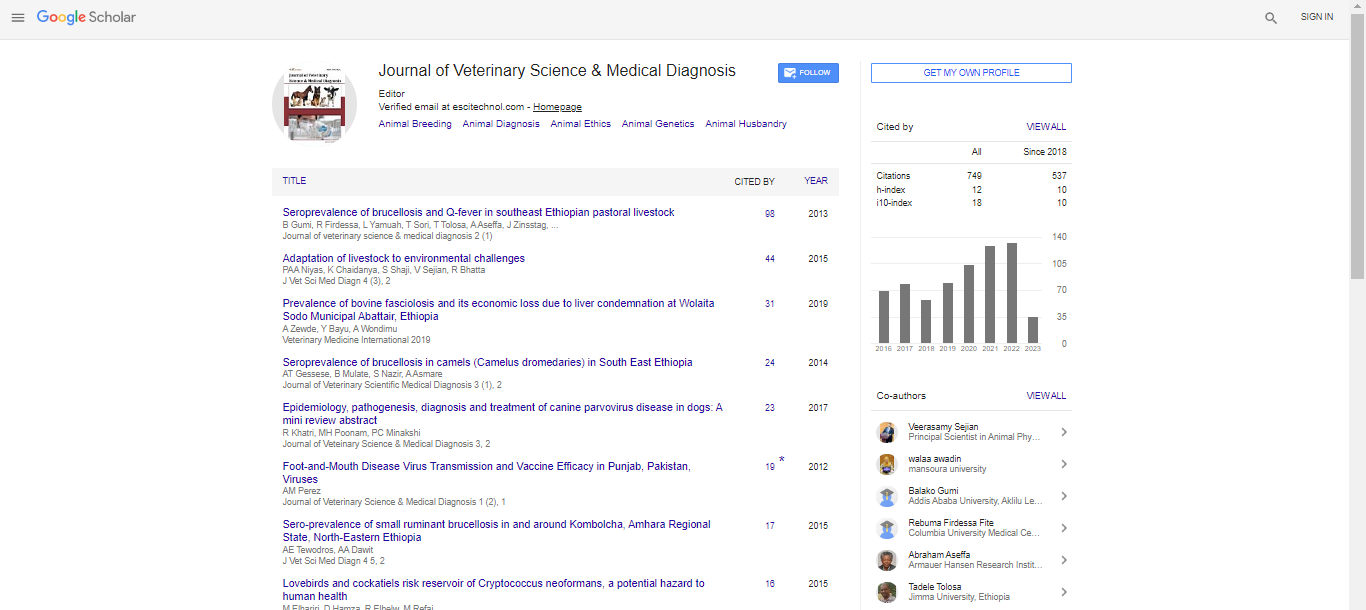
Comparisons between GPX kinetc data in specimens was analyzed, either with or without vitamin E treatment, by student-T (Figure 1,2,3), the significance limit of results is set at P<0.05.

Figure 1: Michaelis and Menten kinetic analysis of glutathione peroxidase activities of RBC membranes from (A) Animals at rest (Rest), (B) Animals at rest after vitamin E diet treatment (Rest Vit E). Each point is the mean (± SEM) of 6 independent determinations. Comparisons are performed by student-T, the significance limit of results is set at p<0,05. For more details, see results

Figure 2: Kinetic analysis of glutathione peroxidase activities of RBC membranes from (A) Trained animals before Cross-Country (C-C pre), (B) Trained animals before Cross-Country vitamin E treatment (C-C pre Vit E).
Each point is the mean (± SEM) of 6 independent determinations. Comparisons are performed by student-T (Figure 2), the significance limit of results is set at p<0.05. For more details, see results.

Figure 3: Kinetic analysis of glutathione peroxidase activities of RBC membranes from (A) Animals after Cross-Country (C-C post), (B) Animals after Cross-Country and vitamin E diet treatment(C-C post Vit E).
Each point is the mean (± SEM) of 6 independent determinations. Comparisons are performed by student-T (Figure 3), the significance limit of results is set at p<0.05. For more details, see results.
Comparisons between fluorescence polarization data are performed by one-way ANOVA and Mann-Whitney post hoc test (Figures 4 and 5) on results from first segment (a) and second segment (b) trends in each plot. There is no statistically significant difference at P<0.05 level.

Figure 4: Fluorescence polarization data (1/P) of purified RBCs from 10 trained horses for each group: (A) At rest; (B) After cross country competition; Data are represented as Perrin plots of 1/P, reciprocal of fluorescence polarization, versus T/h, ratio of absolute temperature on solvent viscosity.
The regression lines are obtained by least square method. Each point is the mean (± SEM) of 10 independent determinations.
The comparisons between data plots by ANOVA are significant for the following P values for both segments (a and b) give P<0.01; both comparisons of A versus B insert and C versus D insert give P<0.01, i.e. are all significant.
The differences among the mean values of the treatment groups are not great enough to exclude the possibility that these are due to random sampling variability at P<0,05 level. For more details, see Results.

Figure 5: Fluorescence polarization data (P) of purified RBCs from 10 trained horses for each group. Data of RBC membranes to which is added vitamin E at 5 mm final concentration (respectively A at rest, and B after exercise).
The statistical analyses were evaluated in graphpad Software Prism® 5.02 program. The regression analysis was performed by IBM SPSS® 17.0.2 program by one-way ANOVA [12,13], according to the least square method. Results are expressed as means ± SE (standard error on the mean).
Results
The GPX activity parameters for animals both before (at rest) and after exercise (cross-country) competitions are referred in (Figures 1-3). The GPX kinetic data depend on Vmax (Table 3) which is evaluated at several GSH substrate concentrations (but at constant cumene hydroperoxide concentration (12 mm) (Figures 1-3).
| Animals groups | Vmax (µM/min/mg prot.) | Apparent Km (µM) |
|---|---|---|
| Rest | 2.72E-05 | 12.25 |
| C-C pre | 2.25E-05 | 9.062 |
| C-C post | 1.97E-05 | 6.85 |
| Rest Vit E | 3.23E-05 | 14.7 |
| C-C pre Vit E | 2.97E-05 | 12.61 |
| C-C post Vit E | 2.59E-05 | 10.53 |
Table 3: Values of GPX Vmax and Km.
To verify the hypothesis that GPX activity is the main scavenger, its activity is also determined under the action of the chemical scavenger vitamin E (50 µM). As described in (Figures 1A, 2A and 3A) and Table 3, the untreated RBCs from rest animals (Figure 1A) show GPX activity Vmax higher than those of trained horses both before (Figure 2A) and after cross-country exercise (Figure 3A). The physiological level (50 µM) of vitamin E in vitro (Figures 1B, 2B and 3B) and Table 3 induces the recovery of GPX activity Vmax after exercise up to levels similar to those of rest animals. Liposoluble reducing vitamin E is highly effective for recovering membrane enzyme activity and its contribution is relevant for all the GPX activities, according to previously described statistical methods.
To confirm the effect of both exercise and vitamin E on the assayed system, fluorescence polarization studies of RBC membranes were performed.
In Figure 4 describes the fluorescence polarization data of RBC membranes of reference animals (Figures 4A and 5A) in comparison with exercised animals (Figures 4B and 5B). The vitamin E treatment (Figure 5B) admits the recovery of fluorescence polarization from oxidative stress up to almost rest levels (Figures 4A and 5A).
The differences among the mean values of the treatment groups are not great enough to exclude the possibility that these are due to random sampling variability; there is no statistically significant difference at P<0.05 level (PaA=0.072) for the first segment (Figure 4A) and (PbA=0.158) for the second one (Figure 4A) (rest animals) using a one-way ANOVA. Similarly for Figure 4B (animals after exercise), Figure 5A (rest animals, vitamin E treated RBC), and Figure 5B (animals after exercise, vitamin E treated RBC) insets of Figures 4,5, the ANOVA analyses respectively give the following pairs of probabilities: (PaB=0.066) (PbB=0.784), (P aC=0.071) (PbC=0.417) and (PaD=0.070) (PbD=0.687).
The results allow ascertaining that the intensive physical activity in the horse produces oxidant radicals, from muscle and respiratory stress, and induces an increase of GPX activity. The addition of α-tocopherol to the incubation buffer of GPX indeed scavenges radicals and relieves this enzyme from exceeding ROSs accumulated long lasting standardized exercise. The fluorescence polarization data show that before the exercise, RBC membranes possess a regular pattern of viscosity but the data from horse blood samples, obtained after the chosen standard exercise (cross-country), show a high viscosity pattern (Figure 4A and 4B). If vitamin E is added to RBC purified membranes the fluorescence polarization is relieved, almost completely flattened also in samples obtained and studied after exercise (Figure 5A and 5B). The obtained results indicate that the training and diet supplements (vitamin E) [7] are able to increase significantly horse antioxidant defences in both the extracellular fluids and blood cells of the tested horses, thus decreasing peroxidative phenomena following physical exercise 3.
The haematological data were determined on blood of both horses underwent to exercise and at rest. Table 2 shows the data of RBC counts (RBC) before and after agonist activity. The Table 2 also sets out the parameters for the Haemoglobin (Hb) and Haematocrit (Ht). From the examination of the described Table 2 follows a high blood content of HB and larger Ht. The described values always increase at the end of the exercise.
Discussion
The results about GPX activity (Figures 1-3) make evident in vitro action of vitamin E on RBC plasma membranes. The oxidative damage induced by standardized physical activity requires effective action by all scavengers recover RBC plasma membrane functions. After the increase of oxidative stress, in fact, the GPX activity damage is evident in exercised horses. The kinetic data in literature show that the GPX enzyme complex activity is probably under the cooperative control of (GSH), as this chemical is both GPX substrate and GRD (Glutathione Reductase) product and is cycled by these enzymes, according to Gallo et al. [9]. At saturating concentrations of GSH, the GPX Km can be calculated (Figures 1A, 2A and 3A). Protective endogenous antioxidant enzymes and exogenous dietary antioxidants help to prevent oxidative damage. Some authors studied other reducing enzymes, but not GPX in horse, that is the most efficient one among athletic mammalians [24]. Only after direct in vitro vitamin E treatment, the GPX activity is comparable to levels before physical activity (Figures 1B, 2B and 3B), confirming the synergy between the vitamin factor and RBCs enzyme scavengers. The action of vitamin E restores the chemical and physical properties of RBC membranes without interference of in vivo metabolic pathways, as also described in membrane fluidity data (Figures 4 and 5).
Optimal levels of membrane fluidity are essential for numerous cell functions including cell growth, solute transport and signal transduction. Exercise produces a dramatic drop in the erythrocyte membrane fluidity as compared to resting time. The highest erythrocyte membrane rigidity was detected after strenuous exercise until exhaustion was performed. Protein carbonyl levels were higher after exhaustive exercises than at rest. Continuous progressive and strenuous exercises until exhaustion, but not submaximal workload, resulted in a significant enhanced accumulation of carbonated proteins in the plasma. These findings are consistent with the idea that exercise exaggerates oxidative damage, which may contribute, at least partially, to explain the rigidity in the membrane of the erythrocytes due to acute exercise [26]. Prior to vitamin E supplementation, exercise induced a significant decrease in EMF or Erythrocyte Membrane Lipid Dynamics (EMLD) vitamin E concentration and a significant increase in MDA concentration at t+1d. Is extremely rapid and does not require 10 weeks of vitamin E supplementation. These results suggested that vitamin E supplementation had a protective effect on submaximal exercise-induced oxidative damage in sedentari dogs. Physical exercise in humans is characterized by an increase in oxygen consumption by the whole body. This leads to a decrease in antioxidant levels that could promote both an increase in the biomarkers and damage to the erythrocyte membrane with consequent modification of membrane fluidity. The sportsmen showed higher levels of the following plasmatic antioxidants: alpha-tocopherol (P=0.03) and superoxide dismutase activity (P=0.0001). According to this evidence, erythrocytes membrane fluidity (P=0.0152) was found to be significantly higher in the soccer players.
The addition of α-tocopherol at almost physiological levels to GPX incubation media confirms that the use of vitamin E partially restores the RBC properties, as described by fluorescence polarization data in preparations from exercised animals (Figure 5A and 5B in comparison to Figure 4A and 4B). Previously [3] studying pentathlon horse blood observed impairments of their biochemical and antioxidant variables (for instance GPX activity, vitamin E levels). Antioxidant adequate food supplementation in healthy horses has neither apparent beneficial nor detrimental effect on pulmonary function in moderate exercise 6, according to Avellini [7,25].
Thus the compensation of RBC number is very pronounced, also considering the presence of RBC stores in these animals, selected and specifically trained by many generations for the described intense physical activities. When an animal is subjected to physical activity, the RBC data show Ht that increases and MCV is almost invariant. For instance, the Hb concentration in blood increases after exercise. This would lead to the hypothesis that the increase in the RBC number involves corpuscular components coming from the squeezing of the spleen, evident after physical exercise, and that more RBCs are released in systemic circulation, containing elements. Such a hypothesis appears validated by the data from the comparison between exercised horses and the ones at rest (Table 2).
Conclusion
The comprehensive examination of the results allows us to describe how the activity of the GPX-GRD complex experiences alterations after physical exercise, in horses. The Km differences of GPX activities are significant between data before and after cross-country exercise. Before exercise, both with and without vitamin E in vitro administration, the GPX activity is comparable with that of resting animals. The GPX data, after exercise, well correlate with the most significant alterations in the RBC population. After physical exercise, an important phenomenon is the presence of polymorphic red blood cells, index of unripe RBCs, coming from other anatomical stores such as bone marrow and spleen. When the reducing vitamin E is added to RBC reaction medium, these cells are almost completely restored to reference functional status. The cross country horses give a small variation of GPX activity in the presence of reducing agents; instead the animals at rest give a larger GPX activity. The experiments with vitamin E and haemolisates, point to the fact that there is formation of reduced GSH from GSSG. The peroxide load in exercised horses is relevant, and the GPX activity decreases according to GPX Km. The in vitro contribution of reducing vitamin E has a relevant role in improving performance in non-worn-out animals, while it is still effective in trained animals, before strenuous effort. The intensive physical activity increases red blood cell count, albeit defective helps to overwhelm the oxidative stress, so that high concentrations of reducing vitamins are partially effective. In described results the extended exercise induces significant derangements of antioxidant GPX activity and decreases the horse antioxidative abilities. ROS main target in the cellular membrane is the lipid fraction. All the described products, as a whole, damage mobility of the membrane phospholipids and diminish the fluidity of the same membranes. The unsaturation’s of membrane phospholipid fatty acids chains should diminish and are converted to radicals or completely oxidized to hydroxilic and aldehyde compounds. When ROSs oxidize enzymes, RBCs are subjected to membrane reorganization which is difficult to reverse and require a long lasting repairing process.
Contributions
GG carried out GPX method optimization and evaluation. GM, director of research, carried out statistical analysis of spectroscopic data. GG Evaluation of haematological parameters. GM performed horse blood sampling and RBC preparations. GM fluorescence spectroscopy and analysis of RBC membranes.
Funding
Support by Italian Ministry of University (IMU) and research ex 60% funding.
Conflicts of Interest
The authors report no declarations of interest.
Ethics Approval
The study was approved by the University of Calabria ethics committee.
Acknowledgements
Authors thank Prof. Domenico Sturino for his language support and technical revision of manuscript.
References
- Finaud J, Lac G, Filaire E (2006) Oxidative stress: Relationship with excise and training. Sports Med 36: 327-358.
- Villani C, Baragli P, Tedeschi D, Sighieri C (2002) Effects of physical exercise and training on plasma GPx and TAS in horse. Conference of the Italian Society of Hippology (SIDI) New acquisitions in the field of hippology, University of Molise, Campobasso.
- Balogh N, Gaál T, Ribiczeyné PS, Petri A (2008) Biochemical and antioxidant changes in plasma and erythrocytes of pentathlon horses before and after exercise. Vet Clin Pathol 30: 214-218.
- Hargreaves BJ, Kronfeld DS, Waldron JN, Lopes MA, Gay LS, et al. (2002) Antioxidant status and muscle cell leakage during endurance exercise. Equine Vet J 34: 116-121.
- Deaton CM, Marlin DJ, Roberts CA, Smith N, Harris PA, et al. (2002) Antioxidant supplementation and pulmonary function at rest and exercise. Equine Vet J Suppl 34: 58-65.
- Williams CA, Carlucci SA (2006) Oral vitamin E supplementation on oxidative stress, vitamin and antioxidant status in intensely exercised horses. Equine Vet J Suppl 38: 617-621.
- Avellini L, Chiaradia E, Gaiti A (1999) Effect of exercise training, selenium and vitamin E on some free radical scavengers in horses (Equus caballus). Comp Biochem Physiol B Biochem Mol Biol 123: 147-154.
- Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 70: 158-169.
- Gallo G, Martino G (2009) Red blood cell glutathione peroxidase activity in female nulligravid and pregnant rats. Reprod Biol Endocrinol 7: 7.
- Rendle DI, Heller J, Hughes KJ, Innocent GT, Durham AE (2009) Stability of common biochemistry analytes in equine blood stored at room temperature. Equine Vet J 41: 428-432.
- Assenza A, Bergero D, Tarantola M, Piccione G, Caola G (2004) Blood serum branched chain amino acids and tryptophan modifications in horses competing in long-distance rides of different length. J Anim Physiol Anim Nutr 88: 172-177.
- Baragli P, Tedeschi D, Gatta D, Martelli F, Sighieri C (2001) Application of a constant blood withdrawal method in equine exercise physiology studies. Equine Vet J 33: 543-546.
- Baragli P, Ducci M, Gatta D, Gazzano A, Sighieri C (2006) Influence of training on plasma adrenaline and noradrenaline kinetics in untrained standard breds. Equine Vet J Suppl 36: 258-261.
- Shinitzky M, Barenholz Y (1978) Fluidity parameters of lipid regions determined by fluorescence polarization. Biochim Biophys Acta 515: 367-394.
- Szmacinski H, Terpetschnig E, Lakowicz, JR (1996) Synthesis and evaluation of ru-complexes as anisotropy probes for protein hydrodynamics and immunoassays of high-molecular-weight antigens. Biophysical Chemistry 62: 109-120.
- Huang X (2003) Fluorescence polarization competition assay: The range of resolvable inhibitor potency is limited by the affinity of the fluorescent ligand. J Biomol Screen 8: 34-38.
- Sivonová M, Waczulíková I, Kilanczyk E, Hrnciarová M, Bryszewska M, et al. (2004) The effect of Pycnogenol on the erythrocyte membrane fluidity. Gen Physiol Biophys 23: 39-51.
- Kawato S, Kinosita K, Ikegomi A (1977) Dynamic structure of lipid bilayers studied by nanosecond fluorescence techniques. Biochemistry 16: 2319-2324.
- Chen LA, Dale RE, Roth S, Brand L (1977) Nanosecond time-dependent fluorescence depolarization of diphenylhexatriene in dimyristoyllecithin vesicles and the determination of "microviscosity. J Biol Chem 252: 2163-2169.
- Lentz H, Barenholz Y, Thompson TE (1976) Fluorescence depolarization studies of phase transitions and fluidity in phospholipid bilayers. 2 Two-component phosphatidylcholine liposomes. Biochemistry 15: 4529-4537.
- Sivonová M, Waczulíková I, Kilanczyk E, Hrnciarová M, Bryszewska M, et al. (2004) The effect of Pycnogenol on the erythrocyte membrane fluidity. Gen Physiol Biophys 23: 39-51.
- Notin C, Vallon L, Desbordes F, Leleu C (2010) Oral supplementation with superoxide dismutase in Standardbred trotters in training: a double-blind placebo-controlled study. Equine Vet J 38: 375-381.
- Avellini L, Silvestrelli M, Gaiti A (1995) Training-induced modifications in some biochemical defences against free radicals in equine erythrocytes. Vet Res Commun. 19: 179-184.
- Berzosa C, Gómez-Trullén EM, Piedrafita E, Cebrián I, Martínez-Ballarín E (2011) Erythrocyte membrane fluidity and indices of plasmatic oxidative damage after acute physical exercise in humans Eur J Appl Physiol 111: 1127-1133.
- Motta S, Letellier C, Ropert M, Motta C, Thiébault JJ (2009) Protecting effect of vitamin E supplementation on submaximal exercise-induced oxidative stress in sedentary dogs as assessed by erythrocyte membrane fluidity and paraoxonase-1 activity. Vet J 181: 288-295.
- Cazzola R, Russo-Volpe S, Cervato G, Cestaro B (2003) Biochemical assessments of oxidative stress, erythrocyte membrane fluidity and antioxidant status in professional soccer players and sedentary controls. Eur J Clin Invest 33: 924-30.