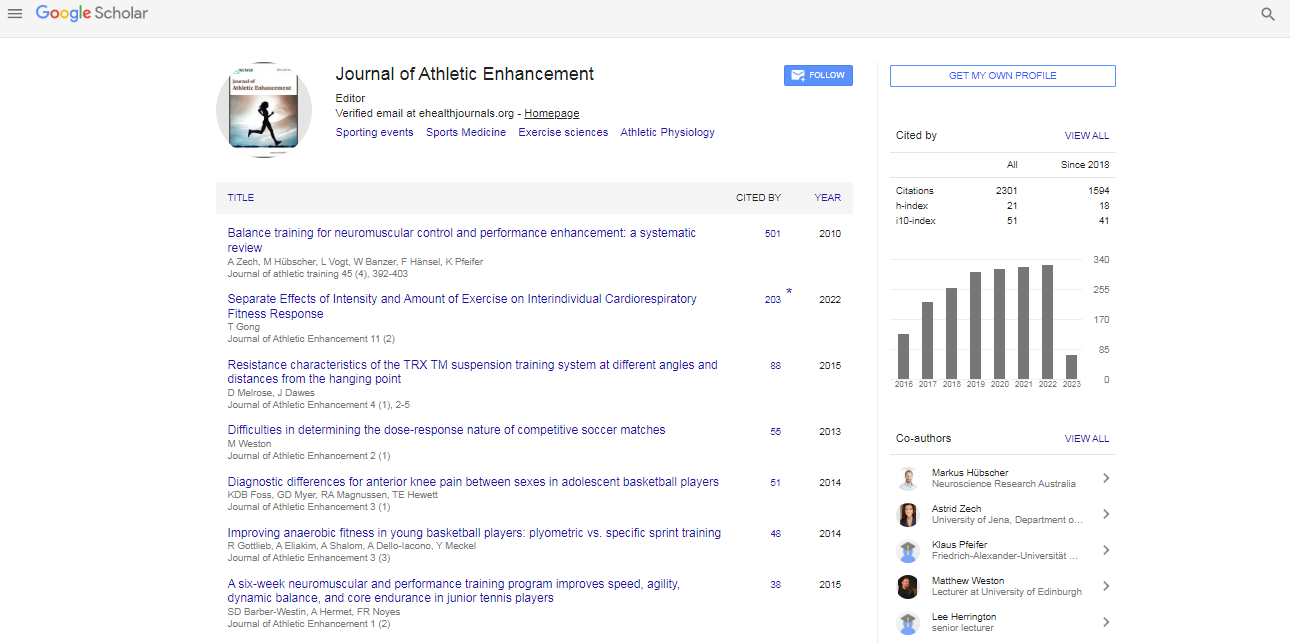
Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, J Athl Enhanc Vol: 8 Issue: 2
The Performance in 10 km Races Depends on Blood Buffering Capacity
Lourenço TF1*, Nunes LAS1, Martins LEB2, Brenzikofer R2 and Macedo DV1
1Laboratory of Exercise Biochemistry (LABEX), Biochemistry Department, Biology Institute, State University of Campinas (UNICAMP), Campinas, Brazil
2Faculty of Physical Education (FEF), State University of Campinas (UNICAMP), Campinas, Brazil
*Corresponding Author : Thiago Fernando Lourenço
Biochemistry Department, Laboratory of Exercise Biochemistry (LABEX), Biology Institute, CP 6109, State University of Campinas (UNICAMP), 13083-970, Campinas, SP, Brazil
Tel: (55) (19) 3521 6146
Fax: (55) (19) 3521 6129
E-mail: thiago.fernando.lourenco@outlook.com
Received: January 21, 2019 Accepted: February 13, 2019 Published: February 20, 2019
Citation: Lourenço TF, Nunes LAS, Martins LEB, Brenzikofer R, Macedo DV (2019) The Performance in 10 km Races Depends on Blood Buffering Capacity. J Athl Enhanc 8:1. doi: 10.4172/2324-9080.1000312
Abstract
For long time running speed related to ventilatory threshold has been considered a strong predictor of running performance ranging from 5 km running to marathon. Specifically for 10 km race, is already known that runners are able to sustain the entire race above it. However, there is no comparison between 10 km performance and running speed related to respiratory compensation point not even about pacing strategy or information regarding acid-base status at this exercise intensity. Objective: We have investigated the relationship between 10 km performance time trial (s10km) and running speed related to ventilatory threshold (sVT) and to respiratory compensation point (sRCP), the pacing strategy related to these intensities and blood acid-base response during four different constant running speeds. Methods: Twelve male amateur (age-37.3 ± 7.2 years; height-171.9 ± 9.4 cm; weight-65.6 ± 10.1 kg; 10 km performance-13.4 ± l1.4 km.h-1) and nineteen elite (age-27.7 ± 9.9 years; height-171.7 ± 7.2 cm; weight-54.7 ± 62.2 kg; 10 km performance-18.6 ± 1.4 km.h-1) long-distance runners performed three exercise protocols: i) 10 km time trial, ii) incremental maximal exercise and iii) four constant load exercise related to sVT, sRCP and above that 72 hours apart. Results: Linear regression analysis revealed that sRCP was the best parameters to predict 10 km runner performance (R2=0.92; p<0.05). No differences was found between the intercept and slope of the relationship between sRCP and s10 km and the identity line (F=0.03; p>0.05) and during almost of entire race, the athletes choose a running speed not different from sRCP. During constant load protocol, all runners completed 10 km at sVT and sRCP, however 3 of 12 amateur and 9 of 19 elite athletes were able to complete 10 km above sRCP. All athletes who completed 10 km in the continuous loading protocols did not show significant changes in blood ph (p>0.05). However, it was not observed in the athletes who were unable to finish. Conclusion: In practice, this study showed sRCP as a useful and safe parameter to predict 10 km running performance and determine pacing strategies for runners, and that the blood buffering capacity is key to maintaining 10 km pacing strategy and performance.
Keywords: 10 km races; Running speed; Linear regression; Strategy; Performance
Introduction
It is already known that endurance performance is dependent on the sum of aerobic and anaerobic metabolism added to biomechanical factors and psychological factors [1]. Physiologically, 10 km running performance depends on the ability to sustain running speeds at high percentages of maximal oxygen consumption (V�?�?O2max) during the main part of the race [1-3]. For this, the knowledge and determination of absolute running speed is crucial to prescribe exercise intensity and determine pacing strategy during these events [4]. However, according to Prud’Homme et al. [5] many studies in this area does not have trustworthy data due unreliable protocols used, which may result in significant over or underestimated results. The gold standard protocol is incremental VO2max protocols with gas exchange analysis to unequivocally determine [6] sub-maximal parameters such as ventilatory threshold (VT) and respiratory compensation point (RCP) [7].
For long time running speed related to VT (sVT) has been considered a strong predictor of running performance ranging from 5 km running to marathon [8,9]. Physiologically, sVT represents a metabolic condition where is observed an increase in anaerobic contribution to energy production [10,11]. Specifically for 10 km race, is already know that runners are able to sustain the entire race above sVT [12-14]. Above sVT, the increased H+ ions removed from the muscle (via the Na+-H+ exchangers and monocarboxylates transporters) may overwhelm the available serum bicarbonate ions (HCO3-) concentrations [15,16]. These higher H+ ions concentrations are mainly detected by central receptors (located in the medulla) and peripheral receptors (located in the carotid body) stimulating the ventilation [17,18]. These exercise intensity is defined as speed related to respiratory compensation point (sRCP) indicating the limit of blood buffering capacity which above that acidosis is installed [11,19]. To best of our knowledge, only one study has related sRCP to 10 km running performance founding no relationship between them [20]. The authors found that VO2max and maximal strength were the main determinants of 10 km running in amateur runners. However, no comparison about pacing strategy and no information regarding acid-base status was investigated in this study.
Recently, Hureau, Romer and Amann [21] strongly proposed that acidosis can stimulate peripheral receptors, via group III/IV muscle afferent, and may induce reductions in central motor drive. In theory, the decreasing in exercise intensity to protect all the body system would influence the runners pacing strategy in 10 km. In fact, this suggestion has been proposed for decades [22], however, there are no studies relating sRCP, pacing strategy and blood acid-base status in field and laboratory conditions may due methodological limitations. Furthermore, there is no information about there is no information regarding whether runners choose running speeds close or not to sRCP during 10 km time trial and how they distribute the exercise intensity relative to it.
With this in mind, our hypothesis is that blood acidosis may limits 10 km running performance independently of training status and sRCP may indicate the exercise intensity chosen by the athletes to complete 10 km without fatigue. To test it we first investigated, in amateur and elite runners, the relationship between 10 km performance time trial and the sVT, sRCP and sVO2max obtained in laboratory conditions through a reliable protocol. Secondly, we investigated blood acid-base response during different constant speeds related to sVT, sRCP and above that in laboratory conditions.
Methods
Subjects
Participated of this study twelve male amateur competing at regional level (A) and nineteen elite long distance runners of the top ten national ranking of 5 km and the 10 km competing in national and international competitions (E) Table 1 presented the physical and metabolic characteristics of the athletes, which confirms two different groups regarding to training status (Table 1).
| A (n=12) | E (n=19) | |
|---|---|---|
| Age (years) | 37.3 ± 7.2 | 27.7 ± 9.9* |
| Height (cm) | 171.9 ± 9.4 | 171.7 ± 7.2 |
| Weight (kg) | 65.6 ± 10.1 | 54.7 ± 6.2 |
| Training experience (years) | 3.3 ± 1.6 | 10.4 ± 4.8* |
| Mean 10 km running speed (km·h-1) | 13.4 ± 1.4Φ | 18.6 ± 1.4Φ* |
| sVT (km·h-1) | 11.5 ± 1.1 | 15.9 ± 1.0* |
| sRCP (km·h-1) | 13.2 ± 1.3Φ | 17.9 ± 1.4Φ* |
| sVO2max(km·h-1) | 16.7 ± 1.2Φ# | 21.0 ± 1.37Φ#* |
| VO2max (ml·kg-1·min-1) | 57.5 ± 9.6 | 74.8 ± 5.2* |
Legend: *-p<0.05 related to A; φ-p<0.05 related to sVT; #-p<0.05 related to sRCP;
sVT-Running speed related to ventilatory threshold; sRCP-Running speed related
to respiratory compensation point; sVO2max- Running speed related to maximal
oxygen consumption.
Table 1: Characteristics of participants. Data are available in mean ± standarderror of mean.
Experimental design
Amateur participants attended the laboratory on six separate occasions and elite runner on three times due their routine schedule. Initially both groups performed individually a 10 km time trial on a 400 m outdoor track recording the time to cover this total distance. After two days they underwent a maximal incremental running test to determine sVT, sRCP and sVO2max. The third test started three days later when the participants randomly performed four 10 km running tests at constant speeds on sVT, sRCP, above sRCP (S1) and at sVO2max with at least 72 hs of rest between them. The S1 was defined as 25% of the difference between sRCP and sVO2max as previously demonstrated by Pires et al. [23]. The E group have made the constant load speed only at S1, due to their schedule limitation. We choose this exercise intensity to coincide with our hypothesis that if blood buffer capacity is, in fact, limiting during 10 km running. The Figure 1 shows the experimental design (Figure 1).

Figure 1: Experimental Design. Legend: VT – Ventilatory Threshold; RCP – Respiratory Compensation Point; VO2max – Maximal oxygen consumption.
Test 1: 10 Km running trial
Ten to 15 minutes warm-up period preceded the test that began at 9 AM. The mean temperature was 24.2 ± 2.2°C and the air humidity was 47.4 ± 1.8% and the runners were allowed to ad libitum hydration during the trial. Each subject was verbally encouraged to give maximum effort and could not use any kind of time device during the running. We recorded every 400 m time to calculate the average running speed of each lap as well as the total test time to determine average speed over 10 km distance (s10 km).
Test 2. Maximal incremental test
In laboratory, after 3 min warm-up at 8-8.5 kmâ�?�?h‾1 the treadmill (Inbrasport Super-ATL, Porto Alegre, RS, Brazil) was set at 9 kmâ�?�?h‾1 and 12 kmâ�?�?h‾1 at a fixed grade of 1% for A and E groups, respectively. These initial running speeds were determined as running speed reached in previous familiarization sessions. After each 25 s interval the speed was increased by 0.3 kmâ�?�?h‾1 until volitional exhaustion [7].
Oxygen uptake (VO2), carbon dioxide output (VCO2), breathing frequency (Bf) and tidal volume (Vt) were continuously collected with an automated breath-by-breath system (CPX/D Med Graphics, St. Paul, MN) using a nafion filter tube and a turbine flow meter (opto-electric). The respiratory exchange ratio (RER) were a quotient of VCO2 on VO2 and to decrease the variability in breath-by-breath acquisition we used the average each 25 s of exercise as recommended by Robergs, Dwyer and Astorino [24]. Prior to each test the analyzer was calibrated using a known gas mixture (12% O2 and 5% CO2), and the volume sensor was calibrated using a 3 L syringe. The laboratory temperature was at 21 ± 1°C and the relative air humidity was between 45-50%.
VT and RCP determinations
The VT and RCP were determined using the V-Slope method from gas exchange measurements [25,26]. The VT detected by the loss of linearity of VCO2 as a function of VO2 during incremental test. According to Whipp and Ward [27] the additional non-metabolic CO2 production resulted from H+ concentration buffering by plasma HCO3 - ions results in a higher change in VCO2 relative to VO2. The RCP was also detected by V-slope method by the loss of linearity of the VE vs. VCO2 relationship [25]. The software supplied by Medical Graphics BreezeSuiteTM 6.4 (MediGraphicsTM) suffered visual inspection by three independent and experienced researchers. The VO2max were considered as the values related to the last completed stage with respiratory exchange ratio (RER) greater than 1.10 [28].
Test 3: Running constant-speed test
Each running constant-speed test (sVT, sRCP, S1 and sVO2max) was preceded by 5 min warm-up at 8-12 kmâ�?�?h‾1. After that, the running speed was set and the protocol was interrupted when the athletes had completed the distance or reached volitional exhaustion. For athletes who did not complete the 10 km we considered the total distance covered until the voluntary exhaustion.
Blood analysis
During each constant-speed test were collected through disposable lancets (Accu-Chek Softclix®, Roche®) and heparinized capillary (Clinitubes®, Radiometer Copenhagen®) approximately 210 μL of capillary blood by fingerstick before (Pre) and at each 2 km intervals. The samples were immediately analyzed in blood gas analyzer Phox Stat Profile (Nova Biomedical®, MA, USA) to obtain blood pH (bpH), blood HCO3 - (bHCO3 -) and blood lactate concentration (bLAC-). The formula of Dill and Costill [29] was applied to all parameters of the blood gas analysis in order to avoid possible interference in their concentrations in response to plasma volume changes induced by exercise.
Statistical analysis
All data are as mean and standard error of mean (mean ± SEM). We tested possible differences between the average speeds during the 10 km (25 laps) and cardiorespiratory parameters by analysis of variance for two factors (two-way ANOVA) with Tukey’s posttest. The relationship between s10km and sVT, sRCP and sVO2max were established by linear regression analysis and determination coefficient (R2) derived from Pearson correlation coefficient. Analysis of covariance (ANCOVA) to compare possible differences between the parameters of the linear regression (i.e. intercept and slope) used the s10km as a covariate analysis. We also used the t-test for possible differences in absolute and relative values of blood parameters between A and E group. The distribution of data obtained from blood were tested by the Shapiro-Wilk test and changes in blood parameters at each time of collection were tested by one-way ANOVA with Tukey's post-test. The level of significance was set <0.05.
Results
Cardio-respiratory measurements and 10 km time trial
As expected, all parameters were significantly higher in E than A group (p<0.05). As shown in Table 1, the sVT was significantly lower and sVO2max was significantly higher than the s10 km (p<0.05) in both groups. Only the comparison between s10 km and sRCP showed no differences between them (p=0.65).
10 km time trial on a 400-m outdoor track and speeds related to maximal incremental test
Figure 2 shows 400-m average running speed over 10 km in comparison with sVT, sRCP, S1 and sVO2max analyses for A and E groups.

Figure 2: Average running speed (± SEM) every 400 m of amateur (A) and elite runners (B) during the 10 km time trial. The horizontal dashed lines represent the mean values of sVT (below), S1 (in the center) and sVO2max (above) obtained in the laboratory. The horizontal solid line represents the mean values of sRCP obtained in the laboratory.
The running speed profile shows a first 400 m corresponding to the highest running speed (A=14.0 ± 1.3 km·h-1; E=18.4 ± 1.1 km·h-1) higher than sVT and sRCP and lower than sVO2max (p<0.05). After that the running speed followed a slight but continuous decline in running speed up to 9.2 to 9.6 km (A=13.4 ± 0.9 km�?�?h-1; E=17.6 ± 1.4 km·h-1). In this period the running speed was not different from sRCP (p<0.05) relative to each group. Then, the running speed of the last 400 m increased significantly (A=13.7 ± 0.9 km·h-1; E=18.3 ± 1.8 km·h-1) characterizing the end spurt.
The linear regression analysis between s10km and sVT (slope=1.01 ± 0.19; R2=0.85; p<0.05), sRCP (slope=1.19 ± 0.19; R2=0.92; p<0.05), S1 (slope=0.76 ± 0.09; R2=0.90; p<0.05) and sVO2max (slope=0.92 ± 0.31; R2=0.52; p<0.05) revealed that sRCP was the best parameters to predict 10 km runner performance (Figure 3). The analysis of ANCOVA corroborated this information, where did not show differences between the intercept and slope of the relationship between sPCR and s10km (F=0.03; p> 0.05) and the identity line. Although high relationships were observed between s10km and sVT and sVO2max, the same pattern was not found. For both, the identity lines were located outside the confidence interval and ANCOVA revealed lower values for the intercept between s10km and sVT (F=398.3; p <0.05) and higher values for sVO2max (F=623.0; p <0.05) when compared to the identity line. Examination of regression residues (data not shown) did not reveal substantial deviations from random errors and also revealed a constant variation, not presenting outliers. The square root values of the mean of the regression residues (in km·h-1) found were 0.95 for sVT, 0.57 for sRCP, and 0.79 for sVO2max, indicating a better determination of s10km by sRCP.

Figure 3: Scatter plot of s10km vs. sVT (A), sRCP (B) and sVO2max (C). Solid lines represent linear regression with their respective confidence intervals (IC; dashed lines), and the dotted line indicates the identity line (slope = 1).
Constant-speed running protocol and blood gas parameters
All amateur runners completed 10 km at sVT and sRCP. However, 3/12 amateur (S1Ac) and 9/19 elite athletes (S1Ec) were able to complete 10 km at S1. The total distance covered by the athletes that were not able to finish 10 km at S1 was 5.22 ± 2.2 km for the amateur (S1Anc) and 4.46 ± 1.1 km for the elite (S1Enc). As expected the total distance covered at sVO2max was lower (1.6 ± 0.3 km; p<0.05) than covered at sVT and sRCP, ranging from 0.4 to 1.5 km for both groups of runners.
Concerning basal values (Pre) of blood parameters no differences between bpH of A group in all studied intensities (sVT=7.46 ± 0.01; sRCP=7.45 ± 0.02; S1=7.48 ± 0.02; sVO2max=7.46 ± 0.01; p>0.05) was found. These values were not different from those found in basal blood of E group (7.49 ± 0.03; p>0.05). Also, we did not detect any significant differences in absolute values of basal blood lactate for A (sVT=1.6 ± 0.6 mmol·L-1; sRCP=1.9 ± 0.3 mmol·L-1; sVO2max=1.9 ± 0.2 mmol·L-1; p>0.05) and E groups (1.6 ± 0.9 mmol·L-1; p>0.05). However, the basal values of bHCO3- of A group (sVT=26.0 ± 1.7 mmol·L-1; sRCP=25.0 ± 3.4 mmol·L-1; S1=25.7 ± 1.6 mmol·L-1; sVO2max=25.8 ± 2.1 mmol·L-1; p>0.05) were significantly lower than those found in E group (28.3 ± 2,0 mmol·L-1; p<0.05). Because of that the acid-base blood results are showed in normalized data (%) in relation to pre-test values in Figure 4, which shows the behavior of bLAC- that indirectly represents the muscular metabolic stress, the bHCO3 - concentrations representing blood buffering capacity and the bpH at 10 km constant loads in sVT, sRCP and sVO2max.

Figure 4: Variation (%) of blood pH (A and B), lactate (C and D) and bicarbonate (E and F) of amateur and elite runners during 10 km run at constant speed relative sVT, sRCP and sVO2max (upper panel) and S1 (lower panel). Legend: bpH - blood pH; bHCO3- - bicarbonate ion; bLAC- - blood lactate; S1Anc- Amateur runner who completed 10 km at S1; S1Ac-Amateur runner who not completed 10 km at S1; S1Enc-Elite runner who completed 10 km at S1; S1Ec - Elite runner who completed 10 km at S1; a - p<0.05 compared to Pre; * - p<0.05 compared to sVT; ψ - p<0.05 compared to sRCP; † - p<0.05 compared to S1Ec ; # - p<0.05 compared to S1Anc.
The 10 km performed at constant sVT and sRCP did not induce significant changes on bpH when compared to Pre (Figure 4A). The bpH were maintained at 7.47 ± 0.02 and 7.45 ± 0.02 during sVT and sRCP exercise, respectively, and the same behavior was found in the athletes who completed 10 km at S1 (S1Ac and S1Ec), presenting bpH at 7.47 ± 0.02 for S1 Ac and 7.52 ± 0.04 for S1Ec (p>0.05; Figure 4B). On the other hand, the athletes who not completed 10 km at sVO2max (7.32 ± 0.01) and S1 (S1Anc=7.32 ± 0.02; S1Enc=7.20 ± 0.04) showed significant decrease in blood pH in the first few kilometers (p<0.05).
As expected, increasing exercise intensities induced crescent improvement in bLAC- after the first two kilometer with posterior values stabilization (sVT=4.61 ± 0.59 mmol·L-1; sRCP=6.52 ± 0.59 mmol·L-1). As shown in Figures 4C and 4D, the variation in bLAC- in athletes who completed 10-m at S1 were similar with those found in sRCP (p>0.05), however, when we analyzed absolute concentration in these groups, we found higher bLAC- concentrations in amateur runners (7.11 ± 0.50 mmol·L-1) compared with elite runners (6.00 ± 0.36 mmol·L-1). The highest concentrations were found at sVO2max (10.42 ± 0.49 mmol·L-1 p<0.05), which indicates an increase of 268,2% (Figure 4C and 4D). These values are similar to found in athletes that do not completed 10 km at S1 (p>0.05). The group S1Anc and S1Enc showed, in average, increments from 1.8 ± 0.23 mmol·L-1, in the rest, to 8.82 ± 0.55 mmol·L-1 after two kilometers (p<0.05) and maintained constant until exhaustion (p>0.05).
During exercise we also found significant decreases in bHCO3- in all constant speed running (Figures 4E and 4F). At sVT and sRCP the blood HCO3 - decreases ~10% (24.0 ± 1.0 mmol·L-1) and ~20% (20.2 ± 1.09 mmol·L-1) after the first two kilometers without differences until the end of the test, being different between them (p<0.05). The same pattern has occurred in athletes who completed 10 km at S1. Slightly lower, but not significant different, values were found in these groups (S1Ac=20.5 ± 2.5 mmol·L-1; S1Ec=24.5 ± 1.04 mmol·L-1; p>0.05). The athletes that did not complete the 10 km at S1nc and sVO2max showed the greatest decrease in bHCO3 - which ranged from 14.0 ± 0.6 mmol·L-1, in sVO2max, to 15.6 ± 1.0mmol·L-1, in S1 Anc.
Discussion
We hypothesed that sRCP could indicate the exercise intensity chosen by the athletes to complete 10 km without fatigue. We also thought that blood acidosis could limit 10 km running performance independently of training status. In fact, we have shown here that sRCP were stronger correlated with s10 km than sVT and sVO2max (Figure 2) and in almost entire 10 km time trial, all athletes choose their running pace near sRCP detected in laboratory conditions.
To our knowledge, this is the first study that made this comparison. Here we have found a strong relationship (R2=0.92) between sRCP and 10 km average running speed, which was higher than previous studies relating sVT and running performance. Jones and Doust [30] found a strong relationship (r=0.81) between sVT and 8 km running, Bellar et al. [31] and Nicholson and Sleivert [32] found strong, but weaker than the present results, correlations lactate threshold and 10 km velocity (r=0.84) in female and (r=0.78) male runners. More recently, Abad et al. [33-35] found that 10 km running time, running economy and peak treadmill velocity explained near 83% of the variance in 10 km running time with a 1.5 min of standard error of the estimate, what is much time in practice. In contrast, here we found that the regression line between sPCR and s10km was almost exactly positioned in the identity line which represents approximately 10 s and one minute for amateur and elite, respectively.
These data reinforce our hypothesis that sRCP could be a good predictor of 10 km running performance. However, it does not fully explain the pacing strategy adopted during the event because they did not choose a constant running speed during entire race. We found that in the first 400 m of 10 km time trial, 67% of the athletes choose running speeds above sRCP (near S1) with a continuous reduction in running speed toward it. Only in the last lap, the end spurt could be found, again near to S1, characterizing an so-called “U shpapped” pacing strategy [36,37]. This data are very similar those found by Lima-Silva et al. [12] who showed that recreational athletes started the 10 km race at a velocity above the average velocity used for the entire race, which was above the onset blood accumulation.
A number of theories try to explain it [14,38,39]. Between them, the teleoantecipatory system and perceive exertion theories propose that athletes build in their brain a template of estimated work rate needed before the exercise begins. According them the neural motor pattern regulated by afferent information from peripheral systems and by individual perceived exertion (i.e., conscious manifestation of the feelings of effort produced by exercise) accounts for the greatest variance of speed during the start phase and perceived exertion [40,41]. Foster et al. [22] have suggested that athletes may monitor their acid-base equilibrium so that critical values are not reached before the total distance has been covered. Theoretically in running speed above sRCP blood pH falls and acidosis is detected by sensory feedback from working skeletal muscle inducing hyperventilation to control blood pH [15,42]. As a consequence the hyperventilation serves to reduce arterial pressure of CO2 that have direct effect on cerebral blood flow and may decrease the arterial oxygenation in frontal cortex reducing/modifying the neural motor drive to protect the system [19].
This results may justify our findings that 93% of athletes choose sRCP and consequently blood pH maintenance for pacing strategy during 10 km time trial with self-running speed adjustment in the beginning and, mainly, in middle phase (400 to 9600 m). However, they do not agree with Bertuzzi et al. [20] who found no relationship between 10 km runner performance and sRCP in recreational runner, maybe due the difference in analyzed groups, once a wide range of sRCP (from 11 to 18 km·h-1) was found in their study, which could not be statistically sufficient to show sRCP influence in 10 km performance.
To confirm our hypothesis that sRCP could be an secure exercise intensity, we tested four different constant-loads, including at sRCP. We found that the bHCO3 - ions decreased almost 21% with no changes in blood pH, indicating that it was efficiently controlled at sRCP. This data suggests that athletes can perform several minutes of exercise at this intensity without fatigue, as found as previously in intensities lower than sRCP [41]. In this way, was interesting that the basal concentration of blood HCO3- significantly higher in elite athletes was not enough for some runners fully sustain the exercise at S1. It can be explained by a possible inefficiency of the enzyme carbonic anhydrase which can be sensitive to a lower zinc intake [42,43] inducing 10% higher ventilatory response (hyperventilation) and lower time to exhaustion in an exercise performed at 70%VO2max.
Interesting data was also found at S1 intensity. Contrary what we initially expected, few runners from both groups were also able to complete 10 km at constant S1 intensity, with no alterations in blood acid-base parameters. These data agree with those found by Pires et al. [23] in healthy cyclists during 30 minutes at constant load related to 25% of the difference between the second lactate threshold and maximal aerobic power output. For them, these results may be due to an integrative, centrally regulated effort model that regulates the body homeostasis based on the remaining exercise time regulated by perceived effort as discussed above related to sRCP. Our data puts the blood buffering capacity as one more piece in this puzzle. The athletes, that completed 10 km at S1, showed maintenance of blood pH despite of a significant decrease in blood HCO3-, which was similar those found in sVT and sRCP (between 10 to 20%). Another possible explanation is the presence of specific adaptations in some athletes, such as a decrease in chemoreceptors sensitivity due to the long-term exposure to altered arterial plasma H+ and pCO2 [44,45], increased strength and resistance of respiratory muscles [46], higher mitochondrial volume and oxidative enzymes [47] trigged by endurance training.
The last possible doubt that can emerge is that S1 could be statistical and/or physiologically, similar to sRCP. However, we ensured that S1 were higher than the typical error for sRCP [7], which cannot discard an artefact of our VO2max protocol or other limitations. Besides this, two other limitations of the study were the lack of dietary and rating of perceived exertion control during the experiment with can be elucidate by futures investigations and explain the capacity to sustain exercise intensities above sRCP.
Conclusion
In practice, this study showed that sRCP is a useful and safe parameter to predict 10 km running performance and determine pacing strategies for runners. The test can be determined using just one maximal or submaximal exercise protocol in the laboratory or even during a 10 km race on a running track. It allows athletes and coaches bring useful information to prescribe exercise intensity during training and competitions. Furthermore, the results presented here incorporate new information regarding the blood buffering capacity as a limiting for 10 km pacing strategy and performance.
References
- Joyner MJ, Coyle EF (2008) Endurance exercise performance: the physiology of champions. J Physiol 586: 35-44.
- Costil DL, Thomason H (1973) Fractional utilization of the aerobic capacity during distance running. Med Sci Sports 5: 248-252.
- Jacobs RA, Rasmussen P, Siebenmann C, Díaz V, Gassmann M, et al. (2011) Determinants of time trial performance and maximal incremental exercise in highly trained endurance athletes. J Appl Physiol 111: 1422-1430.
- Mann T, Patrick R, Lambert MI (2013) Methods of prescribing relative exercise intensityâ�?¯: physiological and practical considerations. Sport Med 43: 613-625.
- Prud’Homme D, Bouchard C, Leblance C, Landry F, Lortie G, et al. (1984) Reliability of assessments of ventilatory thresholds. J Sports Sci 2: 13-24.
- Bentley DJ, Newell J, Bishop D (2007) Incremental exercise test design and analysis: implications for performance diagnostics in endurance athletes. Sport Med 37: 575-586.
- Lourenço TF, Martins LE, Tesutti LS, Brenzikofer R, Macedo DV De (2011) Reproducibility of an incremental treadmill VO(2)max test with gas exchange analysis for runners. J Strength Cond Res 25: 1994-1999.
- McLaughlin JE, Howley ET, Bassett DR, Thompson DL, Fitzhugh EC (2010) Test of the classic model for predicting endurance running performance. Med Sci Sport Exerc 42: 991-997.
- Faude O, Kindermann W, Meyer T (2009) Lactate threshold concepts: how valid are they? Sport Med 39: 469-490.
- Wasserman K, Whipp BJ, Koyl SN, Beaver WL (1973) Anaerobic threshold and respiratory gas exchange during exercise. J Appl Physiol 35: 236-243.
- Wasserman K, Beaver WL, Sun XG, Stringer WW (2011) Arterial H+ regulation during exercise in humans. Respir Physiol Neurobiol. Respir Physiol Neurobiol 178: 191-195.
- Lima-Silva AE, Bertuzzi RCM, Pires FO, Barros RV, Gagliardi JF (2010) Effect of performance level on pacing strategy during a 10-km running race. Eur J Appl Physiol 108: 1045-1053.
- Weston A., Mbambo Z, Myburgh KH (2000) Running economy of African and Caucasian distance runners. Med Sci Sport Exerc 32: 1130-1134.
- Larsen HB (2003) Kenyan dominance in distance running. Comp Biochem Physiol Part A Mol Integr Physiol 136: 161-170.
- Juel C (2008) Regulation of pH in human skeletal muscle: Adaptations to physical activity. Acta Physiol 193: 17-24.
- Meyer T, Faude O, Scharhag J, Urhausen A, Kindermann W (2004) Is lactic acidosis a cause of exercise induced hyperventilation at the respiratory compensation point? Br J Sports Med 38: 622-625.
- Whipp BJ, Davis JA, Wasserman K (1989) Ventilatory control of the “isocapnic buffering” region in rapidly-incremental exercise. Respir Physiol 76: 357-367.
- Lahiri S, Forster RE (2003) CO2/H+ sensing: Peripheral and central chemoreception. Int J Biochem Cell Biol 35: 1413-1435.
- Bhambhani Y, Malik R, Mookerjee S (2007) Cerebral oxygenation declines at exercise intensities above the respiratory compensation threshold. Respir Physiol Neurobiol 156: 196-202.
- Bertuzzi R, Lima-Silva AE, Pires FO, Damasceno MV, Bueno S, et al. (2014) Pacing strategy determinants during a 10-km running time trial. J Strength Cond Res 28: 1688-1696
- Hureau TJ, Romer LM, Amann M (2018) The “sensory tolerance limit”: A hypothetical construct determining exercise performance? Eur J Sport Sci 18: 13-24.
- Foster C, Schrager M, Snyder AC, Thompson NN (1994) Pacing Strategy and Athletic Performance. Sports Med 17: 77-85.
- Pires FO, Noakes TD, Lima-Silva AE, Bertuzzi R, Ugrinowitsch C, et al. (2011) Cardiopulmonary, blood metabolite and rating of perceived exertion responses to constant exercises performed at different intensities until exhaustion. Br J Sports Med 45: 1119-1125.
- Robergs RA, Dwyer D, Astorino T (2010) Recommendations for improved data processing from expired gas analysis indirect calorimetry. Sport Med 40: 95-111.
- Beaver WL, Wasserman K, Whipp BJ (1986) A new method for detecting anaerobic threshold by gas exchange. J Appl Physiol 60: 2020-2027.
- Whipp BJ, Ward SA (1998) Determinants and control of breathing during muscular exercise. Sport Med 32: 199-211.
- Whipp BJ (2007) Physiological mechanisms dissociating pulmonary CO2 and O2 exchange dynamics during exercise in humans. Exp Physiol 92: 347-355.
- Poole DC, Wilkerson DP, Jones AM (2008) Validity of criteria for establishing maximal O2 uptake during ramp exercise tests. Eur J Appl Physiol 102: 403-410.
- Dill DB, Costil DL (1974) Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J Appl Physiol 37: 247-248.
- Jones AM, Doust JH (1998) The validity of the lactate minimum test for determination of the maximal lactate steady state. Med Sci Sport Exerc 30: 1304-1313.
- Bellar D, Judge L, Davis G (2015) Description and predictive value of a novel method for determining the respiratory compensation point using standard scores. J Strength Cond Res 29: 1433-1438.
- Nicholson RM, Sleivert GG (2001) Indices of lactate threshold and their relationship with 10-km running velocity. Med Sci Sport Exerc 33: 339-342.
- Abad CCC, Barros RV, Bertuzzi R, Gagliardi JFL, Lima-Silva AE, et al. (2016) 10 Km running performance predicted by a multiple linear regression model with allometrically adjusted variables. J Hum Kinet 51: 193-200.
- de Koning JJ, Foster C, Bakkum A, Kloppenburg S, Thiel C, et al. (2011) Regulation of pacing strategy during athletic competition. PLoS One 6: 2-7.
- Abbiss CR, Laursen PB (2008) Describing and understanding pacing strategies during athletic competition. Sport Med 38: 239-252.
- Tucker R, Noakes TD (2009) The physiological regulation of pacing strategy during exercise: a critical review. Br J Sports Med 43: e1-e1.
- McCormick A, Meijen C, Marcora S (2015) Psychological determinants of whole-body endurance performance. Sport Med 45: 997-1015.
- Tucker R (2009) The anticipatory regulation of performance: the physiological basis for pacing strategies and the development of a perception-based model for exercise performance. Br J Sports Med 43: 392-400.
- De Morree HM, Marcora SM (2012) Frowning muscle activity and perception of effort during constant-workload cycling. Eur J Appl Physiol 112: 1967-1972.
- Taylor J, Amann M, Duchateau J, Meeusen R, Rice C (2016) Neural contributions to muscle fatigue: from the brain to the muscle and back again. Med Sci Sport Exerc 48: 2294-2306.
- Baron B, Noakes TD, Dekerle J, Moullan F, Robin S, et al. (2008) Why does exercise terminate at the maximal lactate steady state intensity? Br J Sports Med 42: 828-833.
- Lukaski H (2005) Low dietary zinc decreases erythrocyte carbonic anhydrase activities and impairs cardiorespiratory function in men during exercise. Am J Clin Nutr 81: 1045-1051.
- Röcker K, Striegel H, Freund T, Dickhuth HH (1994) Relative functional buffering capacity in 400-meter runners, long-distance runners and untrained individuals. Eur J Appl Physiol Occup Physiol 68: 430-434.
- Dahan A, Nieuwenhuijs D, Teppema L (2007) Plasticity of central chemoreceptors: Effect of bilateral carotid body resection on central CO2 sensitivity. PLoS Med 4: 1195-1204.
- Shea S, Andres L, Shannon D, Banzett R (1993) Ventilatory responses to exercise in humans lacking ventilatory chemosensitivity. J Physiol 468: 623-640.
- McKenzie DC (2012) Respiratory physiology: adaptations to high-level exercise. Br J Sports Med 46: 381-384.
- Laursen PB, Jenkins DG (2002) The scientific basis for high-intensity interval training: optimising training programmes and maximising performance in highly trained endurance athletes. Sport Med 32: 53-73.