Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, Expert Opin Environ Biol Vol: 2 Issue: 3
Impact of Cadmium, Chromium, Cobalt, Lithium and Manganese to the Growth of Fungi and Production of Enzymes
| E Samuel Hartikainen, Annele Hatakka and Mika A Kahkonen* | |
| Department of Food and Environmental Sciences, Division of Microbiology and Biotechnology, P.O. Box 56, Biocenter 1, FI-00014 University of Helsinki,Finland | |
| Corresponding author : Mika Kahkonen Department of Food and Environmental Sciences, Division of Microbiology and Biotechnology, P.O. Box 56, Biocenter 1, FI-00014 University of Helsinki, Finland, Tel: +358-9-19159332; Fax: +358-(0)9-19159301 E-mail: mika.kahkonen@helsinki.fi |
|
| Received: May 28, 2013 Accepted: December 06, 2013 Published: December 11, 2013 | |
| Citation: Hartikainen ES, Hatakka A, Kahkonen MA (2013) Impact of Cadmium, Chromium, Cobalt, Lithium and Manganese to the Growth of Fungi and Production of Enzymes. Expert Opin Environ Biol 2:3. doi:10.4172/2325-9655.1000109 |
Abstract
Impact of Cadmium, Chromium, Cobalt, Lithium and Manganese to the Growth of Fungi and Production of Enzymes
Impacts of metals to the growth and enzyme production of fungi were tested with Cd- (0-10 mg kg–1), Co- (0-100 mg kg–1), Cr- (0-100 mg kg–1), Li- (0-100 mg kg–1) or Mn- (0-400 mg kg–1) containing media with ABTS [2,2’-azino-bis (3-ethylbentzthiazoline-6-sulfonicacid)] as indicator of oxidative enzymes, laccase and/or peroxidases.
Keywords: Metals; Ascomycets; Basidiomycetes; Oxidative enzymes; Fungi
Keywords |
|
| Metals; Ascomycets; Basidiomycetes; Oxidative enzymes; Fungi | |
Highlights |
|
| I) The color reaction with ABTS was seen in all basidiomycetes without or with metals (Cd, Co, Cr, Li, Mn) indicating tested basidiomycetes produced oxidative enzymes. | |
| II) All tested ascomycetes did not show color zone formation with Mn, Cd, Li and without metals indicating that ascomycetes did not produce oxidative enzymes. | |
| III) The growth of A. praecox was tolerant to Mn, Cr and Li. | |
Introduction |
|
| Cadmium (Cd), chromium (Cr), cobolt (Co), lithium (Li) and manganese (Mn) are metals, which are in concern in the polluted environment. These metals have contaminated mining and industrial sites and their surroundings (0.5-4.2 mg Cd/kg soil-1, 18-5750 mg Cr/ kg soil-1, 6-17 mg Co/kg soil-1, 11-71 mg Li/kg soil-1 and 220-10400 mg Mn/kg) [1-6]. Cd, Cr and Li are used in electronic equipments and batteries [7,8]. Co is used in diverse products and materials including batteries, glass and polyester. Cr, Mn and Cd are released into the environment from fertilizers [9,10]. These five metals (Cd, Cr, Co, Li and Mn) are used in large scale globally. | |
| Ligninolytic wood-rotting and litter-decomposing basidiomycetous fungi and also some ascomycetes produce different sets of extracellular oxidative enzymes consisting of peroxidases (lignin, manganese and versatile peroxidase) and laccase, which can depolymerize lignin, cellulose, hemicellulose and starch, and degrade recalcitrant xenobiotic compounds all the way to carbon dioxide [11-17]. White-rot fungi, such as Phanerochaete chrysosporium, Phlebia radiata and Physporinus rivulosus and several others, and litter decomposing basidiomycetous fungi Stropharia spp. and Agrocybe praecox produce lignin degrading enzymes [17-19]. Ascomycetes degrade mainly carbohydrates in soil and litter, but some ascomycetes can also degrade lignin [17,20-22]. Ascomycetes are the dominant fungi in agricultural soils and basidiomycetes are dominant in forest soils. The Ascomycota form the largest number of fungal clades with separate hyphae in the Kingdom Fungi. These clades have been differentiated from the Basidiomycota clade probably before Devon [23,24]. The ascomycetes represent the largest number of fungal species [25-27]. Heavy metals can decrease the ability of fungi to grow and produce extra- and intracellular enzymes and to bioremediate polycyclic aromatic hydrocarbons (PAHs) in the heavy metal contaminated soils [28-31]. Only a little is known about impacts of Cd, Cr, Co, Li and Mn on the growth and oxidative enzyme production of ascomycetous and basidiomycetous fungi. Particularly impact of metals to ascomycetous fungi is very little known. Fungi are crucial in the degradation of detritus and in the cycling of carbon in the soil in well-balanced ecosystems, and therefore their response to metal load in soil is important in soil health globally. | |
| Five ascomycetous and basidiomycetous fungi were tested in the present study. Tested basidiomycetes Phlebia radiata, Pleurotus pulmonarius and Physisporinus rivulosus are wood decaying whiterot fungi found in boreal forests [32,33]. The litter-degrading basidiomycete Agrocybe praecox is a common inhabitant of bark mulch or wood chips [33]. The basidiomycete Stropharia rugosoannulata grows on wood chips, in gardens, and in other cultivated areas [34]. Tested ascomycetes Alternaria sp., Chaetomium sp., Epicoccum sp., Fusarium sp. and Trichoderma harzianum are common litterdegrading microfungi in boreal agricultural and forest soils [35]. | |
| To find out, if metals (Cd, Cr, Co, Li, Mn) have impact on the fungal growth and production of fungal extracellular enzymes, we measured the impact of selected metals (Cd, Cr, Co, Li, Mn) on the growth and enzyme production of five ascomycetous and five basidiomycetous fungi on the contaminated malt extract agar with ABTS [2,2’-azino-bis (3-ethylbentzthiazoline-6-sulfonic acid)] as an indicator. | |
| The aim of the study is to assess impacts of metals (Cd, Cr, Co, Li, Mn) on the growth and the extracellular oxidoreductive enzyme production of five ascomycetous and five basidiomycetous fungi. | |
Materials and Methods |
|
| Fungi | |
| The selected ascomycetous fungi belonging to the genera Alternaria (HAMBI 3289), Chaetomium (HAMBI 3290), Epicoccum (HAMBI 3291) and Fusarium (HAMBI 3292) were identified morphologically on the level of genera by using an optical microscope. These fungi were cultivated from small fragments of hyphae isolated from straw residuals that were buried over winter in arable soil in southern Finland. Buried barley straw residuals were surface sterilized with 70% (v/v) EtOH and 2% (v/v) hypochlorite before placement on growth media containing 2% (w/v) malt extract. Isolated fungi were maintained on 2% (w/v) malt extract agar (MEA) as pure cultures. Pure cultures of identified ascomycetous fungi were stored on 2% (w/v) MEA in sterile glass tubes at +4° C. The four selected ascomycetous fungi are stored in HAMBI culture collection at the Department of Food and Environmental Sciences at University of Helsinki in Finland. Ascomycete Trichoderma harzianum, strain T22, is commercially available fungus, which is commonly used as biocontrol against several fungal soil borne plant pathogens. | |
| The five selected basidiomycetous fungi were obtained from the Fungal Biotechnology Culture Collection (FBCC) at the Department of Food and Environmental Sciences at University of Helsinki in Finland. The basidiomycetes were Agrocybe praecox FBCC 476 (TM70.84), Pleurotus pulmonarius FBCC1465 (ESH1), Phlebia radiata FBCC43, Physisporinus rivulosus FBCC939 (T241i) and Stropharia rugosoannulata FBCC 475 (DSM 11372). They were maintained on 2% (w/v) MEA in sterile glass tubes at +4°C. | |
| Indicator color plate tests | |
| The ability of five ascomycetous and five basidiomycetous fungi to grow and produce extracellular oxidoreductive enzymes was tested with Cd- (0, 2, 5, 10 mg kg–1), Co- (0, 20, 50, 100 mg kg–1), Cr- (0, 20, 50, 100 mg kg–1), Li- (0, 20, 50, 100 mg kg–1) or Mn- (0, 10, 50, 100, 400 mg kg–1) containing media. Used chemicals were Cd(NO3)2•4 H2O, CoCl2•6 H2O, KCr(SO4)2•12 H2O, Li2SO4•H2O and MnSO4•4 H2O. The media contained malt extract agar (MEA, DIFCO) and 250 mg kg–1 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS; Sigma-Aldrich, USA) in the Petri dish plates. All were done as three replicates. Indicator color plate tests were incubated five days at 20 ºC. The diameters of the colored zones (due to the oxidation of ABTS to corresponding cation radical) and the growth of fungi in the plates were measured manually between 90° angels, which the average was calculated in each plate. The intensity of colour was not measured. The formation of the colored zones indicated the fungal extracellular enzymatic reactions. | |
| Statistical tests | |
| To find out statistically significant differences between the growth and enzyme production (color formation zone) of each fungus in the metal (Cd, Co, Cr, Li or Mn) -containing ABTS agar plates compared to the ABTS agar plates without added metal for a certain fungus species (p<0.05), a t-test was done [36]. All statistical tests were performed with Excel (Microsoft) program. | |
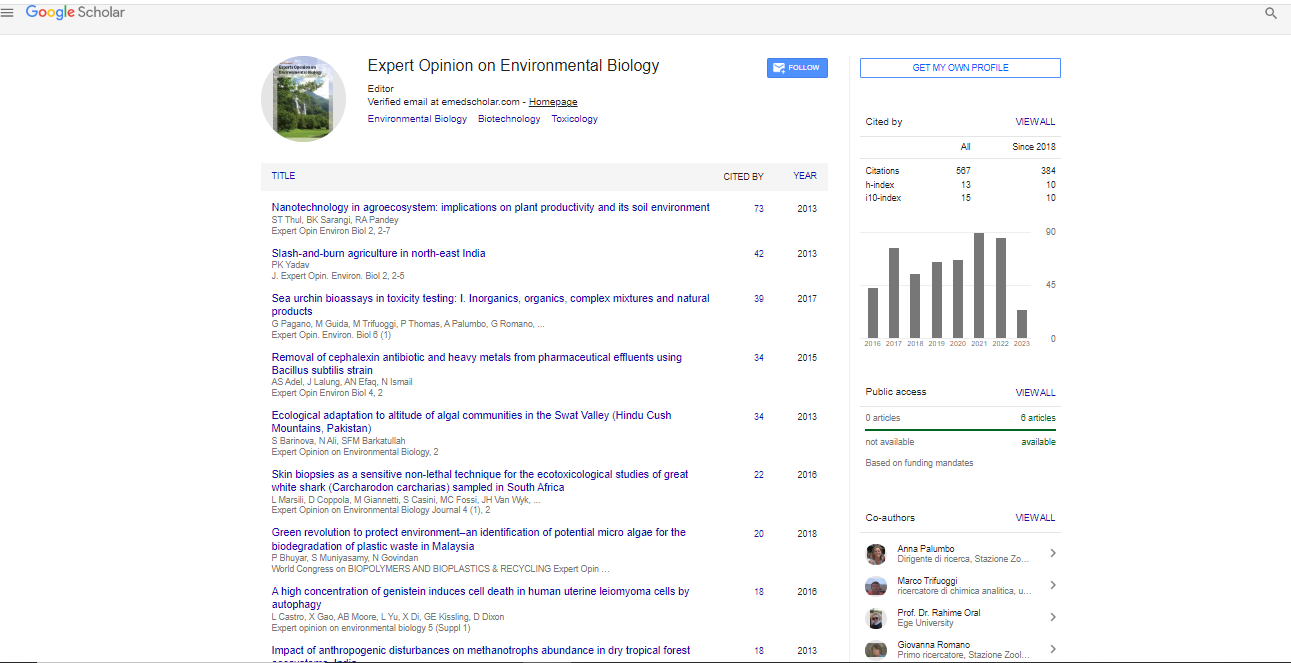
Results |
|
| All five basidiomycetous, A. praecox, P. radiata, P. rivulosus, P. pulmonarius and S. rugosoannulata, fungi grew in the presence of all tested metals (Cd, Cr, Co, Li, Mn) (Figure 1). The results show that the growth of S. rugosoannulata decreased most (45-53%) among five basidiomycetous fungi with 10 mg Mn/kg and with 20- 100 mg Cr/kg compared to the non contaminated plates indicating that S. rugosoannulata was sensitive to Mn and Cr. The growth of P. radiata decreased most among five basidiomycetous fungi with 2 mg Co/kg (63%) and with 100 mg Li/kg (88%). The growth of P. pulmonarius decreased most (100%) among five fungi with 20 mg Co/kg. The results indicate that the growth of the basidiomycetes was most sensitive to Cd (2 mg kg-1) and Li (100 mg kg-1) in the case of P. radiata and to Co (20 mg kg-1) with P. pulmonarius. The growth of A. praecox remained same or even increased with Cr (20-100 mg kg-1), Li (20-100 mg kg-1) and Mn (10-400 mg kg-1) compared to the non contaminated plates indicating that A. praecox was tolerant to Cr, Li and Mn. The growth of P. pulmonarius remained same with Li (20- 100 mg kg-1) and Mn (20-400 mg kg-1) indicating that P. pulmonarius was tolerant to Li and Mn. | |
| Figure 1: Growth of five basidiomycetous fungi in the presence of Mn, Cr, Cd, Co or Li compared to without (100 %) any added metal on ABTS malt extract agar plates (n=3). The error bar indicates standard error of mean. Asterisks (*)=Statistical differences (p<0.05) between with and without added metal. | |
| All five ascomycetes, Alternaria sp., Chaetomium sp., Epicoccum sp., Fusarium sp. and Trichoderma harzianum, grew in the presence of each of the five tested metals (Figure 2). The growth of all tested ascomycetous fungi was similar or even benefitted from Mn (10- 100 mg kg-1) compared to without Mn (Figure 2) indicating that all five ascomycetous fungi tolerated Mn. The growth of Chaetomium sp. among tested ascomycetes decreased most (12%) with 20 mg Cr kg-1, (53%) 2 mg Co kg-1 and (25%) 100 mg Li kg-1. The growth of Alternaria sp. among tested ascomycetes decreased most (60%) with 2 mg Cd kg-1. The results indicate that the growth of tested ascomycetes was most sensitive to Cr, Co and Li in the case of Chaetomium sp. and to Cd with Alternaria sp. The growth of Trichoderma harzianum was similar with Cd (2-10 mg kg-1), Co (20-100 mg kg-1) and Li (20- 100 mg kg-1) compared without metal indicating that Trichoderma harzianum tolerated Cd, Co and Li. | |
| Figure 2: Growth of five ascomycetous fungi in the presence of Mn, Cr, Cd, Co or Li compared to without (100 %) any added metal on ABTS malt extract agar plates (n=3). The error bar indicates standard error of mean. Asterisks (*)=Statistical differences (p<0.05) between with and without added metal. | |
| The color zone formation showing the oxidation of ABTS to cation radical was seen with all five tested basidiomycetes A. praecox, P. radiata, P. rivulosus, P. pulmonarius and S. rugosoannulata fungi with metals (Cd, Cr, Co, Li, Mn) and without tested metals indicating that all five basidiomycetes produced oxidative enzymes (Figure 3). The color zone formation of A. praecox remained stable or even increased with Cd (2-10 mg kg-1), Co (20-100 mg kg-1) and Li (20-100 mg kg-1) compared to the plates without added metals indicating that enzyme production of A. praecox was tolerant and even benefited from Cd, Co and Li. The color zone formation of all five tested basidiomycetes remained stable or even increased with Cr (20-100 mg kg-1) and with Mn (10-50 mg kg-1) compared to the plates without added metals. The color zone formation of P. rivulosus decreased most (40%) among five basidiomycetous fungi with 10 mg Cd kg-1 indicating that P. rivilosus was sensitive to Cd. | |
| Figure 3: Enzyme production (as measured by color zone formation) with five basidiomycetous fungi in the presence of Mn, Cr, Cd, Co or Li compared to without (100 %) any added metal on ABTS malt extract agar plates (n=3). Asterisks (*)=Statistical differences (p<0.05) between with and without added metal. | |
| Ascomycetes did not show any color zone formation in the non contaminated ABTS indicator color plates. There was no color zone formation of any of the five studied ascomycetes, Alternaria sp., Chaetomium sp., Epicoccum sp., Fusarium sp. and Trichoderma harzianum with Mn (10-400 mg Mn kg-1), Cd (2-10 mg Cd kg-1) and Li (20-100 mg Li kg-1). However, color zone formation was observed with added Cr and Co in some cases in ascomycetes plates. Color zone formation of tested ascomycetes was seen with Alternaria sp. with 50-100 mg Cr kg-1 and with Chaetomium sp. and Trichoderma harzianum with 50 mg Cr kg-1. Chaetomium sp. induced color zone formation with 20-100 mg Co/kg. Epicoccum sp. showed color zone formation with 20 and 100 mg Co kg-1. The color zone formation of Alternaria sp. was seen with 100 mg Co kg-1. Color zone formation of tested ascomycetes was not seen with Epicoccum sp. and Fusarium sp. with Cr (20-100 mg kg-1) and with Fusarium sp. and Trichoderma harzianum with Co (20-100 mg kg-1). | |
Discussion |
|
| Our results showed that the growth of the five tested ascomycetous and basidiomycetous fungi had variations between sensitivity to the five tested metals (Mn, Cr, Cd, Li, Co). Impacts of tested metals (Cd, Cr, Co, Li, Mn) on the growth and the production of oxidative enzymes of five basidiomycetes and of five ascomycetes in the present study were not tested in any earlier studies. The growth of the basidiomycetes P. radiata, P. rivulosus, P. pulmonarius and S. rugosoannulata was inhibited in the Cr containing ABTS plates (20-100 mg Cr kg-1) in the present study. These concentrations were 3-25 less than the minimum growth inhibition concentration for Candida sp. isolated from tannery wastes 0.3 g Cr l-1 and for the Rhodosporium sp. isolated from metallurgical wastes 0.5 g Cr l-1 [37]. Phlebia gigantea and Pleurotus ostreatus did not grow with Li (0.5 g l-1) during two weeks in the liquid media [38], which bioavailability of metals is higher than in the solid media in the present study. The contaminant concentration limits set by the Finnish government are 100 mg Co kg-1 soil, 200 mg Cr kg-1 soil and 10 mg Cd kg-1 soil [39]. Finnish Government has not set limit values in the contaminated soil for Mn and Li, which were tested in the present study. The growth of Agaricus bisporus, Agrocybe praecox, Gymnopus peronatus, Gymnopilus sapineus, Mycena galericulata, Gymnopilus luteofolius, Stropharia aeruginosa and Stropharia rugosoannulata was inhibited with 20 mg Ni kg-1 [31] and 22-100% with 200 mg Cu kg-1 [30] on ABTS containing indicator color agar plates. Our results showed that the growth of the most of the tested basidiomycetous fungi was sensitive to Co, Cr and Cd even below the levels of contaminants limits set by the Finnish Authorities. The growth of basidiomycetes P. rivulosus, P. radiata and S. rugosoannulata fungi were vulnerable to Li (100 mg kg-1) and Mn (100 mg kg-1). | |
| The growth of our tested ascomycetous fungi, Alternaria sp., Chaetomium sp., Epicoccum sp. and Fusarium sp., was sensitive to Co, Cr and Cd with concentrations below the soil contaminants limits set by the Finnish government (100 mg Co kg-1 soil, 200 mg Cr kg-1 soil and 10 mg Cd kg-1 soil; [39]. The growth of ascomycetes Chaetomium sp., Epicoccum sp. and Fusarium sp. was sensitive to Li, but limit values have not been set for Li in the contaminated soil. Li is used in batteries and electronic equipments [7,8]. They are mass products and therefore Li is emerging harmful metal in the environment. Trichoderma viride grew 25%, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma pseudokoningii grew 25-50%, Trichoderma harzianum grew 50-75% and Alternaria alternata, Chaetomium globosum, Trichoderma polysporum and Trichoderma reesei grew 75-100% with Li (0.5 g l-1) compared to the control in the liquid media during two weeks [38]. The inhibitory concentrations of Li were 5-25 times higher for ascomycetous fungi than those in the present study for the growth of ascomycetes Chaetomium sp., Epicoccum sp. and Fusarium sp. fungi. Fertilizers contain Cd [10], and this increases the Cd levels of agricultural soils. Finnish agricultural soils are classified as clean if Cd level is less than 0.06 mg kg-1, as normal if Cd level is between 0.06-0.3 mg kg-1 and as unusual if Cd level is more than 0.3 mg Cd kg-1 [10]. Our results indicate that increased level of Cd in the agricultural soil is a threat for the tested ascomycetous fungi, which were isolated from the Finnish agricultural soil. Novel finding in the present study is that the growth of tested ascomycetous fungi was sensitive to Co, Cr, Cd and Li indicating that these metals cause damage and change in the fungal communities in the contaminated soil. These changes can have changes in the cycling of carbon in the contaminated soil. | |
| Ascomycetes did not show color zone formation in the case of all five studied ascomycetes with Mn, Cd and Li and without added metals. Color zone formation was seen with Cr with Alternaria sp., Chaetomium sp. and Trichoderma harzianum and with Co with Alternaria sp., Chaetomium sp. and Epicoccum sp. indicating that Cr and Co induced oxidative enzyme production or at least cation radical formation from ABTS. Cr induced the production of oxidative enzymes with P. rivulosus and three tested ascomycetous fungi. 10-20 mg Cd l-1 has no effect to the activity of laccase with the basidiomycete Pleurotus ostreatus, but laccase activity increases with the increasing metal concentration at 60-600 mg Cd l-1 [40]. Thus taken together, our results show that basidiomycetous fungi were able to produce more oxidative enzymes than ascomycetous fungi with Cd, Cr, Co, Li and Mn and without added metals. Our results indicate that basidiomycetous fungi are more involved in the oxidation of detritus biopolymers with extracellular enzymes in depolymerization process than ascomycetous fungi. Depolymerization of biopolymers is needed in the cycling of nutrients in well balanced ecosystems to maintain soil health globally. | |
Conclusion |
|
| All tested ascomycetes Alternaria sp., Chaetomium sp., Epicoccum sp., Fusarium sp. and Trichoderma harzianum, and basidiomycetes Agrocybe praecox, Pleurotus pulmonarius, Phlebia radiata, Physisporinus rivulosus and Stropharia rugosoannulata, fungi grew with Cd- (0-10 mg kg-1), Co- (0-100 mg kg-1), Cr- (0-100 mg kg-1), Li- (0-100 mg kg-1) or Mn- (0-400 mg kg-1) and without tested metals. The growth of A. praecox was tolerant to Mn, Cr and Li with tested concentrations. The color reaction with ABTS was present in all basidiomycetes without or with metals (Cd, Co, Cr, Li, Mn) indicating tested basidiomycetes produced oxidative enzymes. All tested ascomycetes did not show color zone formation with Mn, Cd, Li and without metals indicating that these fungi did not produce oxidative enzymes. Alternaria sp., Chaetomium sp., Epicoccum sp. and Trichoderma harzianum formed color zone with Co and Cr indicating oxidative enzyme production. Basidiomycetous fungi are more potential to depolymerize detritus biopolymers with oxidative extracellular enzymes than ascomycetous fungi. | |
Acknowledgments |
|
| The study was financially supported by Maj and Tor Nessling Foundation. | |
References |
|
|
|