Spanish
Spanish  Chinese
Chinese  Russian
Russian  German
German  French
French  Japanese
Japanese  Portuguese
Portuguese  Hindi
Hindi Research Article, J Genit Syst Disor Vol: 5 Issue: 2
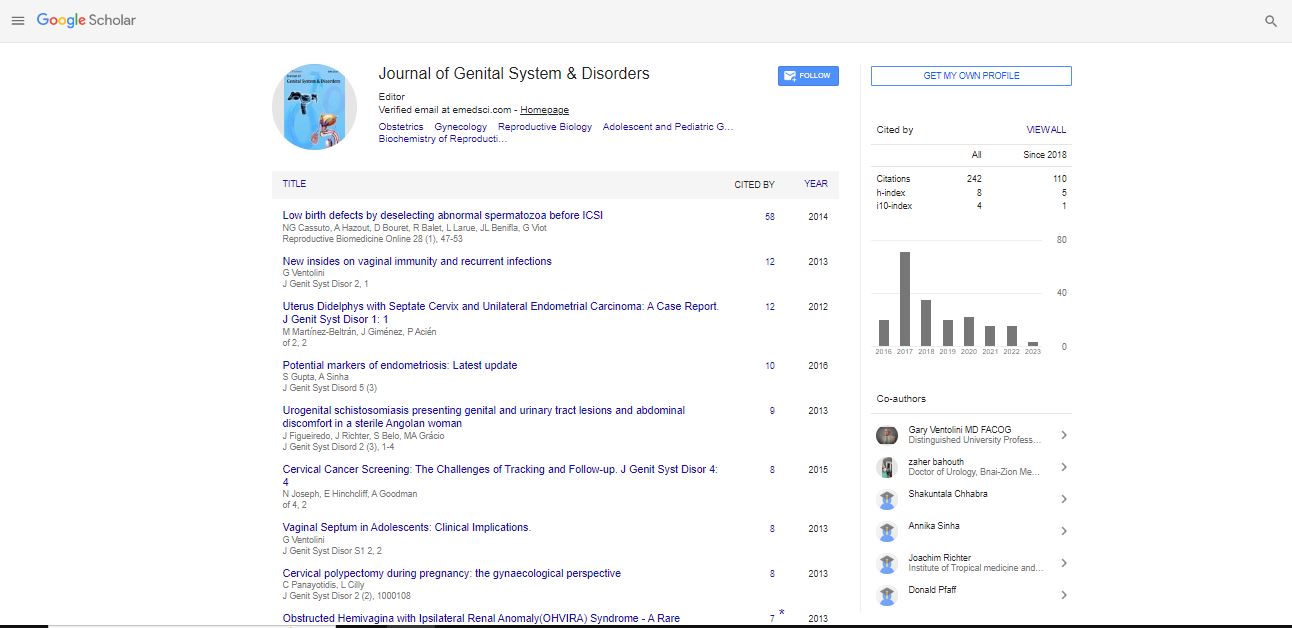
Identification of Merkel Cells in the Labia Minora Skin of Prepubertal Girls
| Schober JM1,4*, Martín-Alguacil N1,2, Cooper RS1,3, Aardsma N3,4, Mayoglou L1,4, Litvin Y1 and Pfaff D1 | |
| 1Department of Neurobiology and Behavior, Rockefeller University, New York, NY, USA | |
| 2Department of Anatomy and Embryology, School of Veterinary Medicine, Universidad Complutense de Madrid, Madrid, Spain | |
| 3Lake Erie College of Osteopathic Medicine, Erie, PA, USA | |
| 4UPMC Hamot, Erie, PA, USA | |
| Corresponding author : Justine Schober, MD 333 State Street, Suite 201, Erie, PA 16507, USA Tel: (814) 455-5900 Fax: (814) 456-0667 E-mail: schobermd@aol.com, voelkerdr@upmc.edu |
|
| Received: February 23, 2016 Accepted: April 12, 2016 Published: April 19, 2016 | |
| Citation: Schober JM, Martín-Alguacil N, Cooper RS, Aardsma N, Mayoglou L, et al. (2016) Identification of Merkel Cells in the Labia Minora Skin of Prepubertal Girls. J Genit Syst Disor 5:2. doi: 10.4172/2325-9728.1000154 |
Abstract
Aim: Merkel cells are located in glabrous skin, hairy skin, and in some mucosa. Clinically significant for their role in sensory perception, neuroendocrine signaling, and MC carcinoma of the vagina, the study of genital tissue sources may enhance understanding of sexual arousal, changes in sexual sensitivity in various hormonal environments, and perhaps even epithelial changes that precede cancer. Details about Merkel cell appearance, location, and function continue to emerge, but little is known about these details in female genital tissue.
Methods: Labial skin samples were obtained from ten normal girls (age 2-9 years) who underwent surgery. The specimens were waste tissue obtained at surgical intervention. All specimen were obtained by the senior investigator, a pediatric urologist, after informed consent had been obtain from the parent or legal guardian. Immunocytochemistry with double immunofluorescence against protein gene product (PGP) 9.5, and CK-20 was performed.
Results: Merkel cells were detected with Merkel cells identified as a single cell isolated near the basal lamina, grouped in the stratum spinosum of the epidermis, around the protrusion of a dermal papilla, or located in more superficial layers of the epidermis in close contact with the axon terminals of sensory nerve fibers. Merkel cells stain by fluorescence immunohistochemistry using antibodies for PGP 9.5 and CK-20.
Conclusion: Merkel cells are identified in the labial minora of prepubertal girls by colocalization with double immunofluorescent staining for PGP 9.5, and CK20.
Keywords: Merkel cell; PGP 9.5; CK 20; DAB; Labia minora
Keywords |
|
| Merkel cell; PGP 9.5; CK 20; DAB; Labia minora | |
Introduction |
|
| Merkel cells were first described by Friedrich Sigmund Merkel in 1875 who observed clusters of cells at the basal epidermis that interacted with nerve fibers. He assumed that these cells performed a sensory function; labeling them “Tastzellen” or “touch cells” [1]. Merkel cells are large oval shaped cells characterized by a lobulated nucleus, dense core vesicles near the nerve fiber junction and microvilli–like projections which contact surrounding keriatinocytes [2] through unique adhering junctions of which desmosomes, composed of plakophilin Pkp2, are most prominent [3]. Merkel cells have been located in a diverse group of mammalian tissues. They have been found in glabrous skin, hairy skin, sinus hairs, ectodermal derived mucosa (oral, anal, and nasal), and taste buds [4,5]. Very little is written about Merkel cell presence and function in female genital tissue, though Merkel cell cancers of the vagina are documented as a rare and often fatal condition. | |
| Immunohistochemical markers have been used to identify Merkel cells in mammalian tissues. Merkel cells have been shown to stain for cytokeratin 8, cytokeratin 18, and cytokeratin 20 [6,7]. Cytokeratin 20 has been shown to be the most specific and sensitive marker used to localize Merkel cells in mammalian tissues [8]. Villin antibodies stain filamental cores found in Merkel cell microvilli-like projections [9]. Merkel cells have also been shown to stain positively for a variety of neuroendocrine markers: met-enkephalin, vasoactive intestinal protein (VIP), substance P, and calcitonin gene-related peptide (CGRP) [10]. Receptors corresponding to neuroendocrine granules have also been shown to be present on the surface of the Merkel cells. The neuron structural protein, protein gene product 9.5 has been used to identify Merkel cells in human oral mucosa [6,11]. Neuronal protein gene product 9.5 (PGP 9.5), is a commonly used marker that is relatively broadly distributed throughout the cytoplasmic compartment, and its immunolabeling has proved to be highly reproducible. | |
| It has been established that Merkel cells are required for proper sensation of light touch [12]. Merkel cells are innervated by slow adapting type 1 tactile nerve, an Aβ-type fiber, which loses its myelin sheath before contacting the Merkel cell. The interaction between slowly adapting type 1 fibers and the Merkel cell is known as the Merkel cell-neurite complex. Slowly adapting type 1 fibers encode sensations such as texture and fine tactile feature discrimination [13,14]. The Merkel cell touch domes are also supplied by Aδ- and C-fibers, which usually conduct nociceptive potentials, leading some to believe the multisensorial nature of the Merkel cell [4,15]. Merkel cells lacking innervation have also been identified further adding to the question of their function. Merkel cells express most of the proteins involved in vesicle trafficking and recycling, they have many components of the glutamatergic transmission machinery, and they bear P/Q-type voltage-gated calcium channels. The latter are normally found in excitable cells, and reveal synaptic capability, since quick calcium currents are believed to be involved in cell depolarization and neurotransmitter release. However, the role of Merkel cells in mechanosensation is still debated. Some say that Merkel cells only direct mechanical stimuli towards the free nerve endings, while others suggest that Merkel cells are involved in mechanotransduction and synapse with the free nerve endings [4,5,13]. | |
| Clinically significant for their role in sensory perception, neuroendocrine signaling, and MC carcinoma of the vagina, the study of genital tissue sources may enhance understanding of sexual arousal, changes in sexual sensitivity in various hormonal environments, and perhaps even epithelial changes that precede cancer. Details about Merkel cell appearance, location, and function continue to emerge, but little is known about these details in female genital tissue. This study seeks to document and characterize Merkel cells within the epithelium of the labia minora. | |
Materials and Methods |
|
| Labial skin samples were obtained from ten normal girls (age 2-9 years) who underwent surgery. The specimens were waste tissue obtained at surgical intervention. All specimen were obtained by the senior investigator, a pediatric urologist, after informed consent had been obtain from the parent or legal guardian. Immunocytochemistry with double immunofluorescence against protein gene product (PGP) 9.5, and CK-20 was performed. | |
| 3,3’-Diaminobenzidine (DAB) Immunocytochemistry | |
| The tissue was post fixed in 4% paraformaldehyde stored in 4 degrees Celsius and cut into 14 micrometer thick sections. Slides were deparaffinized (2x5 min in xylene) and rehydrated (2x3 min 100% ethanol, 1x1 min 90%, 80%, 70% ethanol, 1x5 min de-ionized distilled water). Slides then underwent a process of antigen retrieval in 10 mM citrate buffer in 80 degrees for 20 min. Slices were then washed 3x5 min in PBS, incubated with 1% H2O2 for 20 min, washed three times with 0.1 M PBS (5 min each) and incubated in normal serum (for all antibodies other than CK20: 1% BSA, 1% goat serum 0.2% and triton in PBS; for CK20 1% BSA, horse serum 0.2% and triton in PBS). Slides were incubated overnight at 4�?�?C with one of the following primary antibodies in a humidified chamber: goat anti CK20 (Santa Cruz; sc17113), and rabbit anti PGP 9.5 (abcam; ab53057). Following incubation in primary antibodies, the sections were processed according to a standard procedure for the Vectastain ABC kit (Vector Laboratories). In short, after incubation in the primary antibody, slices were washed three times in 0.1 M anti-goat (for CK 20; vector BA-9500) washed three times in PBS and incubated for 1.5 hr with the avidin-biotin-peroxidase complex in 0.1 M PBS (A and B solutions of the Vectastain ABC kit, Vector Laboratories). Following another set of PBS washes, immunoreactivity was revealed by the addition of the chromogen 3,3’-diaminobenzidine (DAB, 0.02%) to which hydrogen peroxide (0.04%) was added just prior to use and slices were washed three times with 0.1M PBS, dehydrated and cover-slipped. The reaction product appeared as dark brown stain. | |
| Double immunofluorescence | |
| Procedure was identical to DAB staining other than the use of donkey serum. After incubation in primary antibody, slices were washed three times in 0.1 M PBS, and incubated in Cy2 streptavidin donkey anti-goat, Cy3 donkey anti-rabbit (Jackson Immunoresearch) and PBS plus (1% donkey serum and 0.2% triton in PBS) for 1.5 h at a concentration of 1:200. Slides were washed three times in PBS, and incubated for 30 min in 4′,6-Diamidino-2-phenylindole dihydrochloride (DAPI; 1:10000, Sigma), dehydrated and cover slipped. Images were taken using an Inverted LSM 510 laser scanning confocal microscope (Zeiss). | |
| Controls | |
| Controls for primary and secondary antibodies were performed after each round of staining. These were done by omitting the application of one of the four antibodies during the appropriate stage, instead of incubating the tissue with vehicle (PBS plus). We later verified null staining of the omitted antibodies. Also we singlystained tissue using the Diaminobenzidine (DAB) technique and compared our results with each individual stain from the double immunofluorescent staining as far as specificity and location of staining. A PBS plus control was also performed with the DAB single staining. Images were taken using an Inverted LSM 510 laser scanning confocal microscope (Zeiss). Neuronal nuclei expressing levels of fluorescence above tissue background were considered as CK20 and PGP 9.5 positive neurons. | |
Results |
|
| In the labia minora, Merkel cells stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 and CK-20. Merkel cells appear as epithelial cells in close contact with the axon terminals of sensory nerve fibers as shown double labeled (yellow) (Figures 1,3-6). In the labia minora, Merkel cells were detected as a single cell isolated near the basal lamina (Figure 2) or grouped, either forming clusters of cells in the stratum spinosum (Figure 1) around the protrusion of a dermal papilla (Figure 3) or between a dermal protrusion and the basal lamina (Figure 4) when it protrudes through the epidermis. Merkel cells were also located in thinner more superficial layers of the epidermis (Figures 5 and 6). Merkel cells were never located along the whole epidermal layer, but in some spots of the layer. These results were confirmed by DAB staining (Figures 2,3 and 6). | |
| Figure 1: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK- 20 (green). Note the double labeled (yellow) for Merkel cells in closecontact with the axon terminals of sensory nerve fibers. Merkel cells wereidentified isolated near the basal lamina (asterisks) and grouped in the stratum spinosum of the epidermis. 25X. | |
| Figure 2: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK- 20 (green). Cell nuclei counterstain with DAPI (blue). Note the double labeled (yellow) for Merkel cells in close contact with the axon terminals of sensory nerve fibers. Merkel cells were identified in the stratum spinosum of the epidermis. 40x. | |
| Figure 3: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK-20 (green). Note the double labeled (yellow) for Merkel cells in close contact with the axon terminals of sensory nerve fibers. Merkel cells were located around a dermal protrusion within the epidermis. (40X). | |
| Figure 4: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK-20 (green). Note the double labeled (yellow) for Merkel cells in close contact with the axon terminals of sensory nerve fibers. Merkel cells were located within the epidermis, between a dermal protrusion and the basal lamina. (40X). | |
| Figure 5: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK- 20 (green). Cell nuclei counterstain with DAPI (blue). Note the double labeled (yellow) for Merkel cells in close contact with the axon terminals of sensory nerve fibers. Merkel cells were identified near the outer layers of the epidermis in areas where the epidermis was thinner. 40x. | |
| Figure 6: Skin section of the labia minora stained by fluorescence immunohistochemistry using antibodies for PGP 9.5 (red) and CK- 20 (green). Cell nuclei counterstain with DAPI (blue). Note the double labeled (yellow) for Merkel cells in close contact with the axon terminals of sensory nerve fibers. Merkel cells were identified near the outer layers of the epidermis in areas where the epidermis was thinner. 40x. | |
Discussion |
|
| Genital mucousa such as that of the labia minora is recognized as densely innervated, sexually sensitive, influenced hormonally in growth and development, and a site often involved in inflammatory conditions and sometimes epithelial carcinomas. With regards to sensory perception, neuroendocrine signaling, and MC carcinoma, the study of Merkel cells in genital tissue sources may enhance understanding of sexual arousal, changes in sexual sensitivity in various hormonal environments, and perhaps even epithelial changes that precede cancer. | |
| Merkel cells are usually related to epidermal cells scattered in the basal layer of the epidermis and in the outer root sheath of hair follicles. They are characterized by dense core secretory granules and cytoskeletal filaments and synthesize numerous neuropeptides inside the secretory granules. The corresponding receptors are also present at the surface of Merkel cells. The neuropeptide-containing granules are mainly located facing the low-threshold sensory neurons that supply nearly all epidermal Merkel cells. The cluster of Merkel cells with sensory neurons is named the Merkel cell-neurite complex. It constitutes the slowly adapting mechanoreceptor (SAM) reacting in nearby fashion and thus is named type I. Investigations into the exact role of Merkel cells in the perception of touch within the SAM-I have produced conflicting results. Either they are themselves mechanoreceptors, acting as a synaptic transducer to signal sensory neurons, or, rather than the trigger of the neuronal activity, they are the target of sensory neurons in an efferent signal. The synaptic transmission between Merkel cells and neurons has been demonstrated by molecular biology. Merkel cells express most of the proteins involved in vesicle trafficking and recycling, they have many components of the glutamatergic transmission machinery, and they bear P/Q-type voltage-gated calcium channels. The latter are normally found in excitable cells, and reveal synaptic capability, since quick calcium currents are believed to be involved in cell depolarization and neurotransmitter release. Nevertheless, there is still a lack of structural evidence of a synaptic connexion, identification of neurotransmitters and the stimuli that activate Merkel cells. | |
| Immunohistochemical markers have been used to identify Merkel cells in mammalian tissues. Merkel cells have been shown to stain for cytokeratin 8, cytokeratin 18, and cytokeratin 20 [6,7]. Cytokeratin 20 has been shown to be the most specific and sensitive marker used to localize Merkel cells in mammalian tissues [8]. Villin antibodies stain filamental cores found in Merkel cell microvilli-like projections [9]. | |
| In this study, Merkel cells were identified isolated near the basal lamina or grouped as Merkel ending complexes that are disc-like endings flattened against specialized Merkel cells. These clusters were located in the stratum spinosum at the base of the thickened intermediate epidermal ridges. Another location for Merkel cells in the epidermis was around or near the dermal papillae, that project into the undersurface of the epidermis. Merkel discs consist of a nerve terminal and a flattened non-neural epithelial cell. Mechanoreceptive Aβ-neurones are the most represented subset of neurons supplying Merkel cells in SAM-1. Merkel cells are closely associated with the expanded terminal bulb of afferent myelinated nerve fibers. The neuron terminal loses its Schwann cell covering and immediately penetrates the basal lamina, where it expands into a disk or platelike ending that lies in close apposition to the base of the Merkel cell. The combination of the neuron and epidermal cell, called a Merkel neurite complex, is a sensitive mechanoreceptor. However, recent findings show that C and Aδ-fibres also innervate Merkel cells [16], demonstrating that the formation of the SAM is dependent on multiple neurotrophins and their receptors [17]. Each Merkel disc lies adjacent to vesicles within the epithelial cell. The epithelial cell is the mechanically sensitive part, because it makes a synapse-like junction with the nerve terminal. | |
| There is some evidence that light pressure (touch) on the epithelial cell releases from the vesicles a transmitter that stimulates the Merkel disc. The papillae are complemented by what appear to be similar epidermis protrusions, called epidermal ridges, that project into the dermis. If the plane of section is parallel to the surface of the epidermis and passes at a level that includes the dermal papillae, the epidermal tissue appears as a continuous sheet of epithelium, containing circular islands of connective tissue. Merkel cells are thus located at a site where increased mechanical stimuli is placed on the skin, the epidermal ridges are much deeper, and the derma papillae are much longer and more closely spaced, creating a more extensive interface between the dermis and the epidermis. Each of these discs lies adjacent to vesicles within the epithelial cell. The epithelial cell is the mechanically sensitive part because it makes a synapse-like junction with the nerve terminal. Ionic channels, such as osmotic receptor TRPV4 and the purinergic receptors P2Y2, are present on Merkel cells [18]. Swelling-induced hypo-osmolarity may be able to activate Merkel cells through the TRPV4 receptor,while the P2Y2 receptors may mobilize the intracellular calcium required for cell excitability and neuropeptide release. The presence of one stretch activated channel in Merkel cells would support the idea that they are mechanosensory cells. The glutamatergic compon components present in Merkel cells (mGluR5 receptor, subunits of the AMPA and NMDA receptors, VgluT1, 2 and 3) [18-21] reveal their capacity to modulate excitability of neurons, rather than signal transduction. Furthermore, the glutamate receptors are more specific to postsynaptic elements than presynaptic ones [22]. However, they also should be capable of activating sensory neurons of the SAM following their depolarization and the release of their neurosecretory granules, transduced information from touch. Hence, Merkel cells appear to be excitable cells able to transduce stimuli toward several sensory nerve types, and other epidermal cells. They act in touch perception directly or indirectly. | |
| Merkel cells are also thought to play a variety of roles in addition to mechanosensation and mechanotransduction such as neuroendocrine and chemoreception. Acting as a chemoreceptor, Merkel cells respond to histamine by stimulating the TRPV4 receptor expressed on their cell membranes leading to the release of VIP. VIP is known to have immuno-modulatory effects of decreasing proinflammatory cytokines secretion and increasing proliferation of keratinocytes [23]. Merkel cells are found in close relationship with Langerhans cells of the skin and are thought to play a regulatory role in Langerhans cell activation [6]. Merkel cells may also serve a neuroendocrine role regulating vascular, immune, and pigmentary systems [4]. Merkel cells contain a number of dense core granules that contain many different neuropeptides. In vitro studies have shown that VIP and substance P promote growth and differentiation of many cutaneous cells including endothelial and keratinocytes [8]. CGRP, also seen in these dense granules, is known to regulate Langerhans cells. Met-enkaphalin activates granulocyte adherence and migration [4]. In a rat model, receptors to met-enkephalin, VIP, substance P, and CGRP have been localized to Merkel cell membranes indicating a possible paracrine or autocrine reaction to the cytokines released from their dense granules [10]. | |
| All epidermal cells (keratinocytes, melanocytes, Langerhans cells and Merkel cells) express sensor proteins and neuropeptide regulating the neuro-immuno-cutaneous system. Hence they must play a part in the epidermal sensory system [23]. The sensory neurons and skin cells may share a common language with the neuromediators as letters. Thus epidermis can be considered a true sensory tissue where sensor proteins and neuron-like properties enable epidermal cells to participate in the skin surface perception through interactions with nerve fibers. Epidermal cells express many sensor proteins like those found in neurons. These proteins are mainly transmembrane proteins which allow transformation of stimuli like touch, osmotic pressure, temperature or chemical stimulations into biochemical intra-cellular messages [23]. Such neuron-like properties permit the whole epidermis to have sensory functions. | |
| Merkel cells are modified epidermal cells located in the stratum basale. They are excitable multisensorial cells in close contact with sensory nerve endings that can receive almost all environmental stimuli including electromagnetic and ultraviolet radiations, temperature, and humidity. They may transfer the environmental information to oocytes with a multisensorial role that has been attributed to Merkel cells. Melanosomes in Merkel cells may be involved in magnetoreception. Merkel cells are bound to adjoining keratinocytes by desmosomes and contain intermediate filaments in the cytoplasm and dense-cored neurosecretory granules. Merkel and other epidermal cells allow molecular exchanges, thereby modulating the functions of the skin [24]. Merkel cells are most abundant in the skin where sensory perception is acute, historically at the basal lamina. The identification of Merkel cells scattered in outer layers of the epithelium of the labia minora in addition to the basal lamina may speak to the unique sensitivity of this tissue. The study of Merkel cell presence, location, and physiology within the genital epithelium during a variety of developmental stages and clinical conditions may enhance our understanding of physiology and development of genital epithelium, sexual arousal and sensitivity, environmental perturbations of genital tissue, vascular and immunologic function. | |
| The present study is a limited pilot demonstrating the location of Merkel cells within the epithelium of the labia minora. Studies in the fetus, mature and menopausal female may vary, as may different epithelial locations such as: clitoris, labia majora, vagina or perineum. Factors such as pigmentation may also be influential. | |
Conclusion |
|
| Merkel cells are identified in the labial minora of prepubertal girls by colocalization with double immunofluorescent staining for PGP 9.5, and CK20. | |
Conflict of interest |
|
| No conflict of interest. | |
Funding source |
|
| No funding source. | |
Ethical approval |
|
| No approval required. | |
References |
|
|
|